


 Email this Article
Email this Article Print
Print Download (PDF)
Download (PDF)
Abstract 
In
a small part of a cloud forest located at 2700 m in the Eastern
Colombian Andean mountain, where the dominant species is the oak
(Quercus humboldtii), litterfall was determined by means of
12 litter traps. They were placed in two oak canopy trees at
different heights, between 12 m and 22 m. It was found that in adult
oak trees whose biomass of green leaves can vary between 20.52 kg and
27.22 kg, litterfall was presented throughout all the year. The
concentrations of N, P, K, S and Ca were lower in litter rather than
in green leaves, and the Mg concentrations did not vary. Q.
humboldtii presented high concentrations of N (18.2 mg g-1)
and low concentrations of P and K. Nevertheless, these two last
nutrients presented a high retranslocation, which suggests that these
two elements are limited in this ecosystem.
Key words:
Cloud Forests, Canopy, Neotropic, Oaks, Bromeliads, Nutrients, Litterfall.
Resumen
En un relicto de bosque de niebla
ubicado a 2700 m en la cordillera Oriental de los Andes de Colombia,
donde la especie dominante es el roble (Quercus humboldtii),
se determinó la caída de hojarasca por medio de la
postura de 12 colectores, los cuales se pusieron en dos árboles
de dosel de roble a diferentes alturas, entre 12 m y 22 m. Se
encontró que en los árboles adultos de roble cuya
biomasa de hojas verdes puede variar entre 20.52
kg y 27.22 kg se presentó una caída de hojarasca
a lo largo de todo el año. Las
concentraciones de N, P, K, S y Ca fueron menores en la hojarasca que
en las hojas verdes, y las concentraciones Mg no variaron. Q.
humboldtii presentó altas concentraciones de N (18.2 mg
g-1) y bajas concentraciones de P y K, sin embargo, estos
dos últimos nutrientes presentaron una alta retranslocación,
lo que sugiere que estos dos elementos son limitantes en este
ecosistema.
Palabras Claves: Bosques de niebla, dosel, neotropico, roble, bromelias, nutrientes, hojarasca.
Introduction
In the biogeochemical cycle of organic matter and mineral elements, litterfall plays an important role in the relations among soil, vegetation and surrounding environment, constituting one of the essential ecological phenomena in the wooded ecosystems (Vitousek et al. 1995). The litter production is an important process in the tropical forest nutrient cycle (Veneklaas 1995), since the litterfall dynamics to the soil, is one of the most important determinants in the renovation of these forests. Besides it is the most important pathway for the return and transference of carbon and nutrients from the aerial part of vegetal communities towards the surface of the soil (Spain 1984).
A forest constitutes an open system with chemical elements going in and out of it, or moving internally within it. The nutrients remain stored in the vegetation, constituting one of the greatest reserves of nutrients in the ecosystem (Vitousek et al. 1995). Later, the nutrients move until the soil by means of falling leaves and rainwater flows. Another proportion of nutrients is stored in litter and in the other organic matter entering into the forest, from where they are gradually released by its decomposition. The circulation of these nutrients in the ecosystem depends on the organic amount of matter and its rate of decomposition (Sundarapandian & Swamy 1999).
In the case of litter production and availability, this is determined by seasonal fluctuations, which are regulated by biological and climatological processes and factors. However, factors such as topography, edaphic conditions, vegetal species, age, and density of the forest are relevant as well (Hernandez et al. 1992). In any type of forest, the massive litterfall takes place every year in a determined time causing a great accumulation of organic remains at certain times, and producing variations in the dynamics of the nutrient cycling (Jenny 1980).
Due to the threats to the cloud forests present in tropical areas, to their barely short knowledge about them, and the lack of information about their location and importance, the acquisition of new information about the functioning of these tropical mountain forests becomes high-priority. All this with the purpose of obtaining basic tools for the implementation of conservation programs in the long-term. In the case of forests dominated by Q. humboldtii, in spite of their importance, the influence that canopy trees oak litterfall can have in the ecosystem had not been investigated. For this reason, the objective of to calculate the litterfall dynamics by individuals placed on the canopy of Q. Humboldtii, and to quantify the contributions of nutrients that these can make through litter.
Materials and Methods
Study
Area
The project was carried out in the Macanal Reserve located in the eastern Colombian Andean Mountain -South America-, in a town called Bojaca, located 27 km far from Bogota. The study area is a montane cloud forest which is in a precipitous area with steep slopes, and it is located at 2700 m above the sea level. The vegetation presents different levels of human intervention, and this is shown in the fragments of mature forests where mature Oak trees (Q. humboldtii) are the predominant species.
The climatology data of the area is registered by the Acapulco weather station of the "Instituto de Hidrología, Meteorología y Estudios Ambientales (IDEAM)". According to the 13 year-old data analysis (from 1990 to 2002), it has a precipitation with a bimodal behavior with an annual average of 61.5 mm of rainfall; in which the most rainfall months that are March, April, and May in the first term of the year, and October and November in the second term. The average annual temperature is about 13º C, with the highest temperature on May with about 13.4ºC and the minimum on July with about 12.6ºC. The relative annual humidity average is about 91.9% for April, June, and July present the highest percentage with about 93%.
Methods
In a fragment of a conserved forest dominated by oak trees (Quercus humboldtii), a plot of 2125 m2 was made in which four oak canopy trees were chosen. Five platforms were built in the upper part of the tree top at an average height of 20 m and 23 m, with the purpose of having access to different parts of the tree and to the litter traps. The trees were selected under the following criteria: 1) they had to canopy trees; 2) they should allow the ascent with climbing equipment; 3) they had to be in good health; and 4) they had to part of the most mature trees of the fragment.
The monthly contributions of litter were measured through the fall of elements throughout a year. For this, twelve litter traps of 65 cm of diameter and 60 cm of depth were placed in two canopy trees, six in each tree. The litter traps were located in different areas of the top, between 12 m and 22 m. These were checked monthly from November 2004 to October 2005, collecting the material to be dried and weighed. This monthly contribution of litter was evaluated in terms of dry weight, contributed per time and surface. On the other hand, six samples of green leaves and six samples of falling leaves were selected, which were determined the contents of P, Mg, S, N, K, and Ca; these analyses were made in the laboratories of "Corpoica - Tibaitata". The contents of nutrients in litter and green leaves were compared by means of a t test or through the nonparametric test W Mann-Whitney, when the assumptions of normality and homogeneity of variances were not fulfilled; these analyses were made in the Statgraphics program plus 4.0.
The biomass estimate of green leaves of the four chosen trees was obtained by means of the gathering of six branches of different trees with approximate dimensions from 2m x 2m x 2m. All the leaves were taken off these branches in order to be dried and weighed. Subsequently, the number of branches that showed a covering of 2m x 2m x 2m was counted in each tree. With this, an approximation of the number of branches and the quantity of biomass that these held was made. The measurement of the covers of each one of the trees was obtained by direct calculation in m2 -this was convert into hectares for some measures- of the area that projects on the ground the top of each one of the individuals. For the determination of covers, it was used the equation proposed by Rangel (1997).
Results
Litterfall
The trees selected to quantify the biomass of green leaves presented heights that were between 25 m and 26 m, with top covers between 209.3 m2 and 253.98 m2. The amount of biomass of green leaves per tree was calculated between 20.52 kg and 27.22 kg (Table 1).
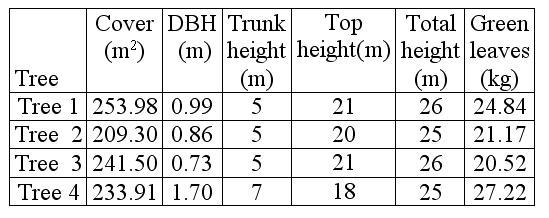
Table 1. Characteristics of the
sampled canopy trees of Quercus humboldtii.In trees 1
and 2, 12 organic matter traps were placed.
Litterfall distribution was uniform in the whole top of both trees throughout all the year, with some variations in the abscission among different months. Nevertheless, the partial results obtained showed that litter availability in this forest occurs throughout all the year. The fall of leaves in tree number one took place throughout all the year, but with greater intensity between the months of November-December-January and June-July; two periods of highest abscission during the months of December and June were registered; in tree number two the litterfall also took place throughout all the year, with greater intensity between January-February and April-May; three periods of highest abscission were registered during the months of January, February and May (Figure 1).
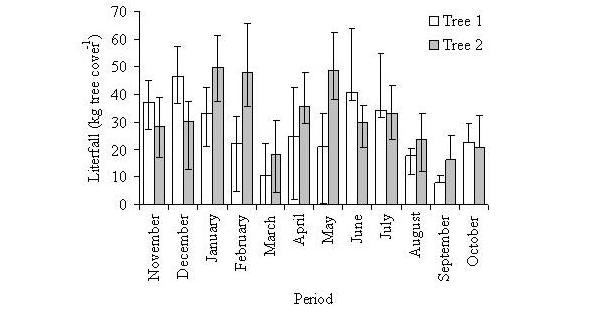
Figure 1. Litterfall (kg to
cover tree-1) at 2700 m, contributed by leaves of canopy
oaks, of trees number 1 and number 2. (Cover of the top: 253.98 m2
and 209.30 m2, trees number 1 and number 2,
respectively).
Nutrients concentration
The concentrations of N, P, K, S and Ca were lower in litter than in green leaves, although these differences were only significant statistically for the content of Potassium and Calcium (W = 3.0, p = 0.02 and t = 2.19, p = 0.05, respectively). The Mg concentrations were similar in fresh leaves and in litter (Table 2).
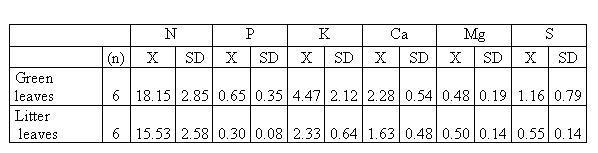
Table 2. Nutrient Contents (mg g-1) in green leaves and litter leaves of the oak (Quercus
humboldtii). Where n is the number of analyzed samples; X is the
average value; SD is the standard deviation.
The approximated annual contributions of nutrients through litterfall from canopy trees were the following: in the tree number one 4.94 kg of N, 0.095 of P, 0.73 of K, 0.51 of Ca, 0.16 of Mg and 0.17 of S for an area of 0.0253 ha corresponding to the cover of the top; in the tree number two 5.92 kg of N, 0.114 of P, 0.88 of K, 0.61 of Ca, 0.19 of Mg and 0.21 of S in an area of 0.0209 ha (Table 3).
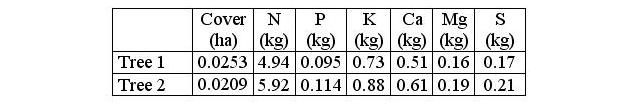
Table 3. Nutrient concentration in (kg) contributed by
each one of the trees throughout a year. Nutrients present in the
fresh leaves of oaks and in the litter expressed in kg.
Discussion
Litterfall
In the nor-western area of the South American Andean range, mainly in Colombia, in the middle part of the mountains, there are important areas of homogenous forests dominated by Q. humboldtii (Lozano & Torres 1974). These forests are covering an altitudinal rank from 1100 m up to 3450 m (Cavelier et al. 2001). In ecosystems dominated by this species, most of the nutrients that enter by means of the fall of organic matter come from litter produced by mature trees of Q. humboldtii. This fact makes the functioning of this ecosystem to be linked to the presence of these trees, as it has been described for other forests; there is a great influence of litterfall of the trees that dominate the canopy in the availability of nutrients in the ground under their tops (Finzi et al. 1998, Saldaña & Lusk 2003). This fact determines the space variation in the availability of nutrients in the ground (Finzi et al. 1998).
In general, mature trees of Q. humboldtii tended to present tops of great cover and great depth, in which an important amount of green leaves all over the top was developed, presenting a high photosynthetic biomass in comparison with other species of canopy trees which grow in other tropical cloud forests, whose values can be between 9.9 kg (Walter & Ataroff 2000) and 15.38 kg (Hofstede et al. 1993) per tree. The depth of the top of these trees and their great amount of biomass of green leaves can be related to the higher tolerance in the shade that this species has in comparison with other arboreal species of cloud forests. It has been registered that the tolerant arboreal species in the shade tend to present tops of greater depth and greater index of foliage area than the intolerants species (Canham et al. 1994).
According to the data collected in relation to the photosynthetic biomass of Q. humboldtii and its litterfall, it was found that foliage renewal in these trees is very high throughout all the year. It is possible that the high litter production can be related to the fact that adult individuals were selected, which might have a more continuous foliage renovation than those individuals in formation stage. This constant litter production of mature Q. Humboldtii probably allows them to be able to keep the nutrients in their leaves until previous moments of their detachment. According to what has been registered, the species with periods of short abscission have determined, with greater precision, the moment in which the leaves are shed. This allows these species to be able to maintain their nutrients until the previous moment to their fall, as well as a high photosynthetic efficiency and a high efficiency in the retranslocation process (Martin et al. 1996).
The greater contributions of organic matter that fell from the top of canopy trees throughout a year were from oak litter. The other organic matter coming from bromeliads, lichens, bryophytes, wood, fruits and flowers neither presented great contributions nor had continuity in their dynamics of fall. Litter production of mature oak canopy trees was high and permanent, reaching a value of 15.4 t ha-1 year-1. In addition to this, the high values of the litter production in this study are also related to the location of the organic matter traps, since these were located at different heights within the top of the same tree from 12 m high, near the places where litter takes place. In most of the studies that are carried out in litterfall, the collectors are set up at a meter from the ground, and part of litterfall that falls is intercepted by epiphytes and another part by vegetation of the intermediate layers and the undergrowth (Veneklaas 1991). Aside from this, the litterfall of trees in its process from fall is often dragged by the wind to distant areas far from where it is placed.
On the other hand, in this investigation the important contributions of biomass that can have mature canopy trees by means of litterfall per hectare is shown. However, the collected data are only generalized about adult individuals of Q. humboldtii, and not about great extensions of cover of this type of forests, since in these forests there is not a continuity of these trees. For this reason, it is possible to incur an overestimate of the aerial biomass and, therefore, of the content of nutrients. Brown and Lugo (1982) affirm that the presence of trees with great diameters can have a great influence on the vegetal biomass as well as on the amount of nutrients.
Leaf nutrients
The chemical analyses of the nutrient contents from leaves and litter of oaks showed that in all the macronutrients except in magnesium, the percentage of concentration is greater in green leaves than in litter. This indicates that before having a process of total fall of leaves, there is a retranslocation of nutrients that is increased in the case of phosphorus and potassium (54 and 48 %, respectively).
When comparing the concentrations of the main nutrients with those of species from other mountain forests of the world, it is observed that the leaves of Q. humboldtii have high concentrations of N (18.2 mg g-1), and its litter contributes a great percentage of this nutrient to the soil of the forest. This can also explain the high productivity of this forest in terms of the production of litterfall and the biomass that the trees of this species reach. The high levels of N in the leaves of oaks have been reported for canopy individuals of Quercus costaricensis and Quercus copeyensis (Kappelle & Leal 1996). These high levels of N in leaves of oaks probably are related to the presence of mycorrhizas in the ground of this type of forests (Chaverri & Rojas 1985)
The concentrations of P were low in comparison with other present vegetal species in other mountain forests of the world (Table 4). Nevertheless, in fresh leaves the levels of P were similar to the reported ones for mountain forests in Puerto Rico (Medina et al. 1981) and in Jamaica (Tanner 1977). In the case of concentrations of K, these were also low in comparison with other vegetal species present in other mountain forests of the world (Table 4), presenting similarity in some values of fresh leaves with the reported ones for mountain forests in Sabah (Proctor et al. 1989) and in Puerto Rico (Medina et al. 1981). The high retranslocation of these two nutrients in oak trees suggests that these elements the availability of these elements is limited in this ecosystem, which has already been suggested for other mountain tropical forests of the world (Vitousek et al. 1995).
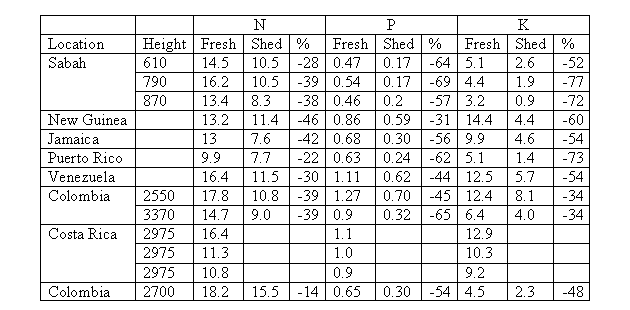
Table 4. Concentrations (mg g-1
dry weight) and percentage of N, P and K in fresh leaves (tree
leaves) and just fallen leaves (litter leaves) in different mountain
forests of the world. References. Sabah: Proctor et
al. (1989); New Guinea: Grubb & Edwards (1982) Jamaica:
Tanner (1977); Venezuela: Steiahard (1979) In: Veneklaas (1991);
Puerto Rico: Medina et al. (1981); Colombia: Veneklaas (1991);
Costa Rica: Kappelle & Leal (1996).
The information provided in this study constitutes a base to emphasize the importance of canopy trees in a forest. In the case of canopy trees of Q. humboldtii, these are an important center for the cycle of several nutrients through their litter contributions, and the nutritional networks that are generated for the return of these nutrients to the ecosystem. At the present time, the presence of these mature trees in fragments of oaks that are still conserved is little, due to the cutting pressure that was exerted on them in previous years. The absence of these trees alters the functioning of these ecosystems, leaving them in certain degree of threat. Apart from this, it should be kept in mind that cloud forests have been investigated superficially in comparison to other tropical ecosystems. However, this is one of the most threatened ecosystems because of the fragmentation and its constant destruction. For this reason there is an important interest in advancing a great number of investigations that allow to know in a detailed way the functioning of these ecosystems with the purpose of being able to take actions to conserve them.
Acknowledgements
This project was carried out thanks to the economic support of the English organization Rufford, Idea Wild support of field equipments and to the logistic support of Fernando Cortes, owner of the Macanal reserve. We thank Sentido Natural Corporation (SN) for the given support through out the project. Especially, we thank Héctor Gasca and Yolima Perez for the document revision. To all the people who participated in the program of SN volunteers for their company to field.
References
Brown, S. & Lugo. A. E. 1982. Storage and production of organic matter in tropical forest and their role in the global carbon cycle. Biotropica. 14, 161-187.
Canhan, C., A. Finzi, S. Pacala & D. Burbank. 1994. Causes and consequences of resources heterogeneity in forest: interspecific variation in light transmission by canopy trees. Canadian Journal of forest Research. 24: 337-349
Cavelier, J., D. Lizcaíno & M.T. Pulido. 2001. Colombia. In: Kapelle M. & A. Brown 2001.Bosques nublados del neotropico. Primera edición
Chaverri, A. & I. Rojas 1985. Ensayo de inoculación de plantulas de robla copey Quercus copeyensis (Mueller) con suelo micorrícico en condiciones de invernadero. In: Ciclo sobre técnicas de investigación en micorrizas. Pp 111-130. International Science Foundation & Catie. Turrialba.
Hernández, I. M., I. Santa-Regina, J.F. Gallardo. 1992. Dinámica de la descomposición forestal en bosques de la cuenca del Duero (Provincia de Zamora): Modelización de la perdida de peso. Soil Research and Rehabilitation. 6: 339-355
Hofstede, R., J. Wolf, & D. Benzing. 1993. Epiphytic Biomass and Nutrient Status of Colombia Upper Montane Rain Forest. Selbyana. 14: 37-45.
Finzi, A., N. VanBreemen & C. Canham 1998. Canopy tree soil interactions within temperate forest: species effects on pH and cations. Ecological Application 8: 447-454
Grubb, P.J. & P.J. Edwars. 1982. Studies of mineral cycling in montane rain forest in New Guinea III. The distribution of mineral elements in above-ground material. Journal Ecology. 70: 623-648.
Jenny, H. 1980. Soil genesis with ecological perspectives. Spinger - Verlag. New York 560 pp.
Kappelle, M. & M. Leal. 1996. Changes in leaf morphology and foliar nutrient status along successional gradient in a Costa Rica Upper Montane Quercus forest. Biotropica 28(3): 331-344
Lozano, G. & J. H. Torres. 1974. Aspectos generales sobre la distribución, sistemática Fitosociológica y clasificación ecológica
Martín, A., I. Santa-Regina & J.F. Gallardo. 1992. Eficiencia, retraslocación y balance de nutrientes en bosques de Quercus pyrenaica bajo diferente pluviometría en la Sierra de Gata (Centro-Oeste Español) Ecología. 10: 79-93
Medina, E. C. & P. Weaver. 1981. Composition foliar y transpiración de especies leñosas de Pico Este, Sierra de Luquillo, Puerto Rico. Acta cient. Venez. 32: 159-165.
Proctor, J., C. Phillips, C.K. Duff, A. Heaney & E.M. Robertson. 1989. Ecological studies on Gunung Silam, a small ultrabasic mountain in Sabah, Malasia II Some forest processes. Journal of Ecology. 77:317-331.
Rangel, O. 1997. Colombia Diversidad Biotica II. 89-381. Universidad Nacional de Colombia & Instituto de Hidrología, Meteorología y Estudios Ambientales IDEAM.
Saldaña A. & C. Lusk. 2003. Influencia de las especies del dosel en la disponibilidad de recursos y regeneración avanzada en un bosque templado lluvioso del sur de chile. Rev. chi. hist. nat. v 76 n 4 Santiago.
Spain, A.V. 1984. Litterfall and the standing crops of litter in three tropical Australian rain forest. Journal of Ecology. 72: 947-961
Sundarapandian, S.M. & P.S. Swamy P.S. 1999. Litter production and leaf-litter decomposition of selected tree species in tropical forest at kodayar in the western Ghats, India. Forest Ecology and Management. 123: 231-244.
Tanner, E.V. J. 1977. Four montane rain forest of Jamaica: a quantitative characterization on floristics the soil and the foliar mineral levels and discussion of the interrelations. Journal Ecology. 65: 883-918.
Veneklaas, E. J. 1991. Litter and nutrient fluxes in two montane tropical rain forest, Colombia. Journal of Tropical Ecology. 7:319-336
Veneklaas, E. J. 1995. Water and nutrient in two montane rain forest canopies, central cordillera, Colombia. Chapter 14. In: Studies on Tropical Andean Ecosystmes. Volume 4. Ecoandes.
Vitousek P., G. Gerrish, D.R. Turner, L. Walker & D. Mueller-Dombois. 1995. Litterfall and nutrient cycling in four Hawaiia Montane rain forest. Journal of Tropical Ecology 11:189-203
Walker R. & M. Ataroff. 2002. Biomasa epífita y su contenido de nutrientes en una selva nublada andina venezolana. Ecotropicos 15(2)