


 Email this Article
Email this Article Print
Print Download (PDF)
Download (PDF)
Resumen 
La investigación se realizó en base de 235 entradas de las diferentes especies de Vasconcelleas colectadas en las provincias de Loja, Zamora Chinchipe, El Oro y parte del Azuay y luego caracterizadas morfológicamente . Para el estudio de cruzabilidad se realizaron 1 337 cruzamientos entre las especies colectadas y se ejecutaron en las dos vías cuando era posible (cruzamiento directo y recíproco); del total de cruzamientos realizados, se obtuvieron 725 frutos cuajados (54.15 %). Los cruzamientos en los que Vasconcellea stipulata Badillo, interviene tanto como madre y padre frente a las otras especies, el cuajado es del 59 %; al cruzar Vasconcellea x heilbornii Badillo como madre, el cuajado es del 53.56 % y como padre 62.22 %; cuando Vasconcellea pubescens Lenné et Koch, interviene como padre, el porcentaje de cuajado es del 68.45 %, pero como madre desciende al 25 %; similar situación se presenta en Vasconcellea candicans Gray como padre con 56.93 % y como madre 25% . Un alto porcentaje de cruzabilidad se da en Vasconcellea x heilbornii Badillo nm. pentágona (heilborn), como madre (70.13%) y Vasconcellea sp. con un 90.01 %, mientras que como padre decrece a un 25 %; finalmente los cruzamientos en Vasconcellea parviflora (A.DC.) Solms. tanto como madre y padre presentan porcentajes bajos: 33.3 % y 34.93 %, respectivamente, al igual que Vasconcellea monoica Desf., con 30 y 23.94 %. El alto nivel de cruzabilidad sugiere que estamos en presencia de una sola especie, con varias subespecies.
Palabras clave: caracterización, cruzamiento, cuajados, especie, subespecie
Abstract
This paper presents the results from crussing 235 samples of different species of Vasconcelleas collected in the provinces of Loja, Zamora Chinchipe, El Oro and part of Azuay and then morphologically characterized. For the cross breeding study 1337 crossings between the collected species were undertaken in two ways when it was possible (direct and reciprocal crossing).
725 of the crossings formed fruits. In the cross breeding Vasconcellea stipulata Badillo works like a mother and father in relation to other species,
with 59 %; of the crosisngs forming fruits. When crossing Vasconcellea x heilbornii Badillo as a mother, 53.6 % formed fruits and as a father 62.22 %. When Vasconcellea pubescens Lenné
et Koch was used as a father, the percentage of formed fruits was 68.45%, but as a mother it decreased to 25 %, similarly to Vasconcellea candicans Gray., as a father with 56.93 % and as a mother 25%. A high percentage of cross breeding is seen in Vasconcellea x heilbornii Badillo nm. pentagona (heilborn), as a mother (70.13 %) and Vasconcellea sp. with 90 %, while as a father it decreases to 25 %; finally the cross breedings in Vasconcellea parviflora (A.DC.) Solms., behaving as a mother and father present low percentages: 33.3% and 34.93 %, respectively, the same as with Vasconcellea monoica Desf., with 30 % and 23.94 %. The high level of cross breeding that we are in presence of only one specie, with several subspecies.
Key words: characterization, cross-breedings, formed fruits, species, subspecies
Introducción
Ecuador, debido a su ubicación geográfica y a su excepcional topografía, contiene un alto porcentaje de la biodiversidad mundial (26 % de la flora nativa) y dentro de ella, la Región Sur del país alberga aproximadamente el 44.91% del total nacional, distribuidos el 19.1 % en la Provincia de Loja, el 17.7 % en Zamora Chinchipe y el 8.1 % en El Oro (Aguirre et al. 2002 cita a Jorgensen y León-Yánez 1999 y a Valencia et al. 2000); parte de ella la constituyen la familia de las Vasconcelleas, cuyas especies algunas de importancia económica regional, encierran un enorme potencial tanto para la selección y mejora de las especies, como para enfrentar los diversos problemas, especialmente de adaptación, respuesta a virosis y calidad de los frutos, entre ellas se cuentan: V. pubescens, V. stipulata. V. candicans, V. x heilbornii, V. parviflora, V. monoica y V. pentagona (Vasconcellea x heilbornii), híbrido natural este último, cultivado tanto en Sudamérica como en Nueva Zelanda, Australia, Israel, Italia, California, Grecia y Brasil, cuyas formas cultivadas proceden del Ecuador y sur de Colombia (Badillo 1999).
Su importancia también ha tomado fuerza últimamente debido a que las hojas contienen alcaloides amargos (carpaína y pseudocarpaína) que actúan como digitalizadores en el corazón y la respiración; adicionalmente contienen dos alcaloides (piperideinos, dehidrocarpaína I y II) que son más potentes que la carpaína. El látex de la planta y frutos verdes contienen dos enzimas proteolíticas, (papaína y chimopapaína); esta última en cantidades más abundantes, siendo la papaína doblemente potente (Proexant 2005), la cual tiene amplias aplicaciones en medicina, con la extracción de la papaína (Joan Sisa 2005), una de las enzimas más versátiles por tener químicamente gran afinidad con el cuerpo humano, así constituye una enzima digestiva por excelencia, muy apreciada por sus atributos de recubrir el estómago y facilitar la digestión al romper las cadenas protéicas entre otros beneficios, por lo que se la usa en medicina para las insuficiencias gástricas, heridas y gangrenas, para disolver membranas en difteria y reducir la hinchazón, fiebre y adhesiones postcirugía; además en la industria culinaria como materia prima para la elaboración de ablandadores de carnes, en la cosmetología para lociones de la piel, en la textilera para suavizar la lana y la seda, para la curtiembre en la maceración de cueros, en la industria cervecera para clarificarla disolviendo las proteínas que la opacan, en la fabricación de chicles y para la preparación de medios de cultivo en laboratorios.(Proexant 2005).
La carencia de germoplasma identificado como fuente de resistencia a enfermedades, principal problema en el cultivo de estas plantas, ha creado la necesidad de explorar alternativas, como las hibridaciones interespecíficas, misma que se basa en el hecho de haber sido registrados híbridos de este tipo en forma natural o artificial, como es el caso de los babacos, también por el hecho de encontrarse resistencia a varias enfermedades en estos parientes (Drew R. et al. 1998); en anteriores experiencias de hibridaciones (Vegas et al. 2003) y otras seis especies del género, se encontró que las barreras precigóticas son mínimas, con una adecuada penetración de los tubos polínicos, las barreras postcigóticas son mayores debido a la aborción de óvulos y endospermo, en este caso, el rescate de embriones es una herramienta valiosa para eliminar esas barreras (Mekako H, et al. 1975). La ventaja con las hibridaciones interespecíficas, radica especialmente en la posibilidad de acceso, por parte de los países de la región a los recursos genéticos, por otra parte, ésta es una resistencia que ha sido consistente en el campo por mucho tiempo y su uso no está sujeto a restricciones y controles, se necesita entonces de materiales genéticos que aseguren una resistencia durable al alcance de nuestros países y con los mínimos costos sociales, económicos y ambientales.
Debido a la aceleración de la erosión genética de estos materiales, el Departamento de Biotecnología de la Universidad Nacional de Loja, hace varios años ha venido realizando colectas, contando en la actualidad, con un Banco Vivo, como una primera medida para asegurar el aprovechamiento sostenible de los recursos genéticos de las Vasconcelleas, los cuales una vez caracterizados y evaluados (Morales R.,et al. 2004), han sido incluidos en programas de mejora, donde se realizan ensayos de cruzamientos, obteniéndose hasta el momento, datos muy interesantes y con características genéticas de interés, con un total de 1 337 cruzas entre las diferentes especies.
La continuación de la colecta, caracterización, cruzabilidad y evaluación del germoplasma aportará, amplias perspectivas en términos de una mejora duradera y sostenible de las Vasconcelleas, especialmente, en los aspectos de adaptabilidad, patología y entomología, lo mismo que para ofrecer al mercado las calidades, sanidad, sabor y apariencia exigidos, en busca de una mayor competitividad en el marco nacional, regional y mundial, para lo cual, el germoplasma aportará la base genética requerida.
Materiales y Métodos
Trabajo previo
Sobre la base del trabajo de un banco de germoplasma con 235 entradas de plantas del género Vasconcellea de la familia Caricaceae (Morales, R. et al. 2004) colectadas en diferentes sitios de las provincias de Loja, Zamora Chinchipe, El Oro y parte del Azuay, mismas que fueron caracterizadas morfológicamente utilizando descriptores creados para el efecto que incluyó 150 variables, se las clasificó taxonómicamente y se determinó su distribución geográfica así como los sitios de mayor variabilidad, finalmente se realizó la caracterización molecular seleccionando 29 entradas de las 235, cuyos resultados muestran sus relaciones filogenéticas entre las diferentes entradas. Las especies que participaron en el presente trabajo son: V. pubescens, V. stipulata, V. candicans, , V. x heilbornii, V. parviflora, V. monoica, V. x heilbornii var. pentagona y Vasconcellea sp., ésta última se la nombra así debido a que las plantas poseen características morfológicas distantes a las demás especies (probablemente una nueva especie).
Cruzamientos
La siguiente fase, se la desarrolló en el presente estudio, en el que se realizaron las hibridaciones entre todas las especies establecidas en el banco vivo entre los meses de septiembre a noviembre del 2005, para lo cual, en las especies que funcionaron como madre las flores se encontraban aún con los pétalos cerrados (en madurez fisiológica) y con la ayuda de una pinza se abrieron las estructuras florales, hasta dejar al descubierto el pistilo, el cual inmediatamente fue fecundado con el polen maduro extraído de la especie que funcionaría como padre, para finalmente colocar la etiqueta en la que se identifica el código de la madre y el del padre, tomando registro de cada uno de los cruzamientos realizados. Dicho procedimiento se lo realizó tomando muy en cuenta las condiciones climáticas (temperatura baja y no lluvioso), puesto que las altas temperaturas reducen considerablemente la tasa de cuajado.
El plan de cruzamiento establecido, contempla la realización de hibridaciones en las dos vías en, la que cada especie funcione como madre y como padre cada vez que era posible, puesto que en los huertos donde se cultiva la planta solamente se encuentran hembras ya que los machos son desechados, por lo que fue necesario utilizar plantas de semilla para encontrar los machos requeridos.
Transcurridas dos semanas se realizó la primera evaluación del porcentaje de cuajamiento y finalmente luego de dos semanas más se grabaron los frutos con el código de sus progenitores.
La evaluación del total de frutos cruzados se la tomó directamente en el campo al momento de realizar cada cruza. La evaluación final de los frutos cuajados se la realizó cinco a seis semanas post hibridación, tomando datos planta por planta.
Finalmente con los datos de campo se realizaron los porcentajes de cuajado y se determinó la cruzabilidad entre las diferentes especies de Vasconcelleas, así como la caracterización de algunos frutos que llegaron a su madurez.
Material vegetal
Códigos:Ecu |
Género |
Especies |
|
|
9645,9655,9654,9637,9739,9654,9663,9646, |
Vasconcellea |
heilbornii |
var.pentagona | |
9645,9661,9662,9634,9643,9655. |
|
|||
9754,9758. |
Vasconcellea |
parviflora |
|
|
9818. |
Vasconcellea |
monoica |
|
|
9632,9764,9790. |
Vasconcellea |
pubescens |
|
|
9765,9860,9765,9861,9752,9630,9783. |
Vasconcellea |
sp. |
|
|
9674,9738,9679,9670,9648,9770,9746,9647, |
|
|||
9775,9733,9738,9633. |
Vasconcellea |
stipulata |
|
|
9709. |
Vasconcellea |
candicans |
|
|
9822-107,9822-108,9822-60,9822-61,9822-62, |
|
|||
9822-110,9822-56,9822-57,9822-53,9822-105, |
|
|||
9822-54,9822-55,9828,9822-49,9822-11,9727, |
|
|||
9822-46,9822-47,9822-98,9822-104,9822-103, |
|
|||
9650,9822-40,9822-42.9822-43,9822-36,9659, |
|
|||
9822-37,822-38,9822-39,9728,9782,9822-120, |
|
|||
9822-112,9822-91,9822-92,9822-89,9822-113, |
|
|||
9822-34,9822-114,9822-115,9822-32,9822-82, |
|
|||
9822-3,9659,9822-87,9822-86,9727,9822-118, |
|
|||
9822-28,9822-116,9822-117,9822-25,9822-27, |
|
|||
9822-26,9828,9666,9822-63,9822-29,9822-35, |
|
|||
9822-64,9822-119,9822-121,9822-99,9822-30, |
|
|||
9822-70,9822-71,9822-72,9781,9828,9822-1, |
|
|||
9836,9822-74,9740,9822-78,9730,9650,9827, |
|
|||
9772,9769,9769,9653,9635,9822,9660,9821, |
|
|||
9650,9772,9781,9730,9728,9781,9666,9653, |
|
|||
9659,9660,9786,9822-97,9822-98,9740,9782, |
|
|||
9822-102,9822-101,9822-59,9822-51,9822-52, |
|
|||
9822-65,9822-68,9822-77,9822-79,9781,9822-73. |
Vasconcellea |
heilbornii |
|
Resultados y Discusión
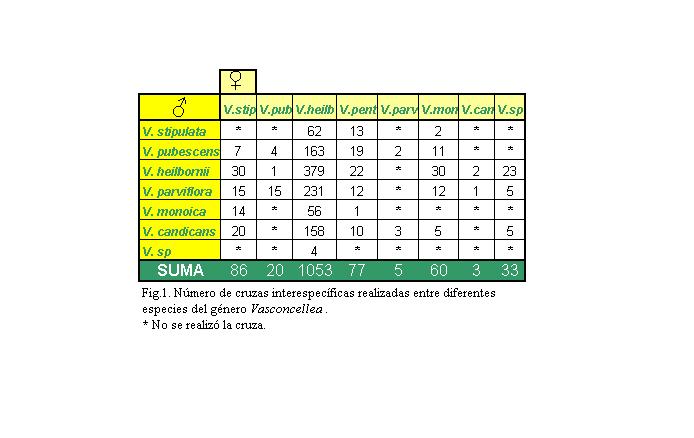
Figura 1: Número
de cruzas interespecíficas realizadas entre diferentes
especies del género Vasconcellea.
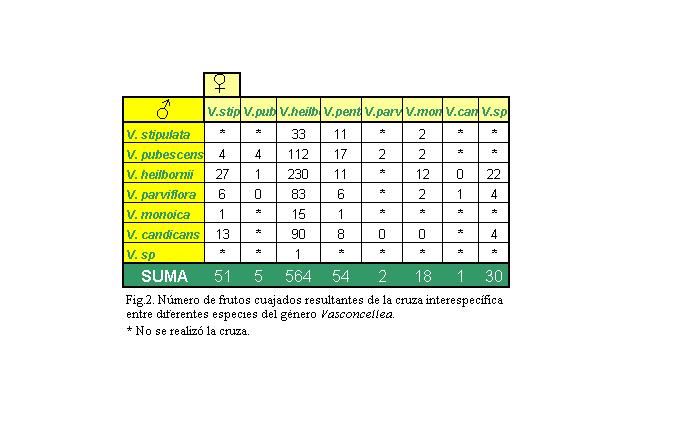
Figura 2: Número
de frutos cuajados resultantes de la cruza interespecífica
entre diferentes especies del género Vasconcelllea.
Cruzamientos de Vasconcellea stipulata como
madre
La tasa media de cuajado a nivel general en todas las especies está entre el 40 y 57 %, observándose que los cruzamientos en los que interviene V. stipulata como madre logra cruzarse con V. parviflora y V. pubescens, en porcentaje medio (40 y 57.14 %), mientras que con V. heilbornii y V. candicans es más alto (90 y 65 %), finalmente con V. monoica presenta gran dificultad de cruzamiento, presentándose solamente en un 7.17 %.
Cruzamientos de Vasconcellea stipulata como padre.
Como padre la cruza de V. stipulata con V. heilbornii está dentro del promedio (53.23 %), mientras que con V. monoica y V. pentágona este porcentaje es muy alto (100 % y 84.62 %), respectivamente.
Cruzamientos de Vasconcellea pubescens como
madre
Las hibridaciones realizadas en V. pubescens como madre, produjeron frutos con V. heilbornii en un 100 % mientras que con V. parviflora no funcionaron.
Cruzamientos de Vasconcellea pubescens como
padre
Las hibridaciones de V. pubescens como padre cuajaron con V. stipulata dentro de la media (57.14 %), con V. heilbornii, V. pentágona y V. parviflora cuajaron en un alto porcentaje (68.71, 89.47 y 100 %), respectivamente, mientras que con V. monoica fue muy bajo de sólo el 18.18 %.
Los frutos caracterizados, producto de la cruza entre (V x heilbornii) x V. pubescens, presentaron tamaño mediano con un aroma que va de suave a fuerte, todos con la piel mediana a lisa y verde amarillenta y una porción comestible alrededor de 77 g.
La cruza entre V. pentagona x V. pubescens dio frutos grandes, de aroma suave, piel mediana a lisa y color amarillo verdoso.
Cruzamientos de Vasconcellea x heilbornii
como madre
Los cruzamientos de V. x heilbornii como madre, se ubicaron bajo la tasa media de cuajado en especies como V. parviflora, V. monoica y Vasconcellea sp. con 35.93, 26.79 y 25 % respectivamente, dentro de la media las especies V. stipulata y V. candicans (53 y 56.96 %) y finalmente en un alto porcentaje V. pubescens con 68.71 % y V. heilbornii con 60.69 %.
Cruzamientos de Vasconcellea x heilbornii
como padre
Los cruzamientos de V x heilbornii como padre se ubicaron en la media con V. pentágona y V. monoica (50 y 40 %), subiendo el promedio con V. stipulata, V. pubescens, V. heilbornii y Vasconcellea sp. (90, 100, 60.69 y 95.65 %), respectivamente, mientras que con V. candicans fue nulo.
Los frutos producidos por la cruza entre (V. x heilbornii) x (V. x heilbornii) dio frutos amarillos verdosos, de aroma suave, piel mediana a lisa y con una porción comestible de 74.25 g.
Cruzamientos de Vasconcellea x heilbornii var.
pentagona como madre
Los cruzamientos de V x heilbornii var. pentagona como madre, produjeron frutos con todas las especies que se cruzó, presentando porcentajes medios con V. heilbornii y V. parviflora (50 %), y mayores porcentajes con V. stipulata, V. pubescens, V. monoica y V. candicans (84.62, 89.47, 100 y 80 %), respectivamente.
Los frutos híbridos de V. x heilbornii var. pentagona x V. parviflora son de color amarillo verdoso, piel mediana a lisa y de aroma fuerte.
La cruza entre V. x heilbornii var. pentagona x V. pubescens produjo frutos grandes, de aroma suave, piel mediana a lisa y color amarillo verdoso.
Cruzamientos de Vasconcellea parviflora como
madre
Los cruzamientos de V. parviflora como madre con V. pubescens funcionaron en un 100 %, mientras que con V. candicans fue nulo.
Cruzamientos de Vasconcellea parviflora como
padre
Los cruzamientos de V. parviflora como padre con V. candicans y Vasconcellea sp. son muy altos (100 y 80 %), en la media se ubicaron los cruzamientos con V. stipulata y V. pentagona (40 y 50 %); con V. heilbornii y V. monoica resultaron muy bajos (35.93 y 16.66 %), mientras que con V. pubescens no se lograron frutos cuajados.
Cruzamientos de Vasconcellea monoica como madre
Los cruzamientos de V. monoica como madre dieron tasas altas de cuajamiento: con V. stipulata (100 %), mientras que cruzo muy difícilmente con V. pubescens, V. heilbornii y V. parviflora (18.18, 40 y 16.66 %) y resultó nula con V. candicans.
Cruzamientos de Vasconcellea monoica como padre
Los cruzamientos con V. monoica como padre con V. stipulata y V. heilbornii son bajos (7.14 y 26.79 %), subiendo al 100 % con V. pentágona.
Cruzamientos de Vasconcellea candicans como
madre
Los cruzamientos de V. candicans como madre con V. parviflora cuajaron en un 100 %, mientras que con V. heilbornii no cuajó ninguno.
Cruzamientos de Vasconcellea candicans como
padre
Los cruzamientos de V. candicans como padre presentaron cifras superiores al promedio, así con V. pentágona, V. stipulata y Vasconcellea sp. (80, 65 y 80 %), con V. heilbornii están dentro de la media con un 56.96 % y finalmente con V. parviflora y V. monoica resultó imposible obtener frutos cuajados.
Cruzamientos de Vasconcellea sp. como madre
Finalmente V. sp., como madre cruzó con V. heilbornii, V. parviflora y V. candicans con una tasa de 95.65, 80 y 80 %, respectivamente.
Cruzamientos de Vasconcellea sp. como padre
La cruza de Vasconcellea sp. como padre con V. heilbornii, tuvo una tasa baja de 25 %.
Conclusiones
Para los cruzamientos de V. candicans
Como padre cruza con V. stipulata, V. x heilbornii, V. x heilbornii var. pentagona y Vasconcellea sp. y como madre solo cruza con. V. parviflora.
Para los cruzamientos de V. parviflora
Como padre cruza con V. stipulata, V. x heilbornii, V. x heilbornii var. pentagona, V. monoica, V. candicans y Vasconcellea sp y como madre solamente cruza con V. pubescens.
Para los cruzamientos de V. monoica
Como padre cruza con V. stipulata, V. x heilbornii y con V. x heilbornii var. pentagona, y como madre cruza con V. pubescens, V. stipulata, V. x heilbornii, y con V. parviflora.
Para los cruzamientos de V. stipulata
Como padre cruza con V. x heilbornii , V. x heilbornii var. pentagona y V. monoica; y como madre cruza con V. pubescens, V. monoica, V. x heilbornii , V. parviflora y V. candicans.
Para los cruzamientos de V. pubescens
Como padre cruza con V. x heilbornii , V. stipulata, V. x heilbornii var. pentagona, V. monoica y V. parviflora; y como madre cruza con: V. x heilbornii .
Para los cruzamientos de V. x heilbornii.
Como padre cruza con : V. stipulata, V. pubescens, V. monoica, V. x heilbornii var. pentagona y Vasconcellea sp. y como madre cruza con V. pubescens, V. monoica, Vasconcellea sp , V. stipulata, V. parviflora y V. candicans.
Para los cruzamientos de V. x heilbornii var. pentagona
Como madre cruza con V. pubescens, V. stipulata, V. monoica, V. parviflora, V. candicans y V. x heilbornii.
Para los cruzamientos de Vasconcellea sp.
Como padre cruza con V. x heilbornii, como madre cruza con: V. x heilbornii, V. parviflora y V. candicans.
La baja tasa de cruzabilidad que presentan V. monoica, V. parviflora y V. candicans corrobora lo mencionado en un artículo anterior (Morales R., et al. 2004), donde se señala que la distancia genética existente entre éstas especies silvestres y las cultivadas, es grande, mientras que estas últimas presentan una estrecha similitud genética (V. stipulata, V. pubescens, V. x heilbornii y V. x heilbornii var. pentagona) entre si.
Referencias
Aguirre, Z. et al. 2002. Botánica austroecuatoriana, estudios sobre los recursos vegetales en las provincias de El Oro, Loja y Zamora Chinchipe. Quito, Ec., Herbario Loja. ABYA YALA.
Badillo, V. 1999. Familia Caricaceae. 2da edición. Universidad Central de Venezuela. Facultad de Agronomía. Maracay, Venezuela. p. 36-101.
Proexant. 2005. El cultivo de la papaya. Producción de exportaciones agrícolas no tradiconales. [email protected]
Joan, Sisa. 2004. Papaya (Carica papaya L.). Ecoaldea.com. (www.ecoaldea.com/plmd/papaya.htm)
Drew, R.; O'Brien, C. &Magdalita P. 1998. Development of interspecific Carica hybrids. Acta Horticult.
Vegas, A.; Trujillo, G. & Sandrea. Y. 2003. Obtención, regeneración y evaluación de híbridos intergenéricos entre Carica papaya Y Vasconcellea cauliflora. INCI. [online]. dic. 2003, vol.28, no.12 , p.710-714.
Mekako, H.; Nakasone, H. 1975. Floral development and compatibility studies of Carica species. J. Amer. Soc. Hort. Sci.
Morales, R.; Medina, D. & Yaguache, B. 2004. Diversidad genética filogenia y distribución geográfica del género Vasconcellea en el Sur del Ecuador. Lyonia.