


 Email this Article
Email this Article Print
Print Download (PDF)
Download (PDF)
Resumen 
El número de esporas por esporangio en el
80% de los helechos (Polypodiales) es 64; no obstante, este número
es variable en Hymenophyllaceae. Al realizar un conteo de las esporas
de 17 especies de Hymenophyllaceae en Costa Rica, se encontró
que el número de esporas por esporangio variaba en un rango de
24 a 240. El carácter tuvo poco significancia taxonómica
a nivel genérico, sin permitir distinción entre
Trichomanes
e Hymenophyllum,
al igual que sus subgrupos. Fortuitamente, se logró
encontrar una diferencia clara entre dos especies cercanamente
emparentadas, H.
fucoides y H.
tunbridgense.
Palabras
Claves: Esporas, Esporangios, Hymenophyllaceae,
Hymenophyllum
Trichomanes.
Abstract
The number of spores per sporangium in 80% of the ferns
(Polypodiales) is 64; however, this number is variable in
Hymenophyllaceae. I collected 17 species of Hymenophyllaceae in Costa
Rica in order to count their spores. The number of spores per
sporangium varied within a range of 24 to 240. The character resulted
of little taxonomic significance at generic level, not allowing
distinction between Trichomanes
and Hymenophyllum,
nor of their sub-groups. One clear difference was unexpectedly found
between two closely related species, H.
tunbridgense fucoides
and H. tunbridgense.
Key words:
Spores, Sporangia, Hymenophyllaceae, Hymenophyllum,
Trichomanes.
Introducción
La espora es la célula reproductiva asexual de un helecho, resultado de un proceso de división celular (meiosis) producido en el esporangio, partiendo de 16 megaesporocitos o células madres diploides (2n), que al dividirse dará lugar a 4 células hijas haploides (n) (Morán, 2004).
Dentro del grupo de los helechos Eusporangiados o helechos verdaderos el número de esporas por esporangio generalmente es múltiplos de cuatro (16, 32, 64, 128, 256 y 512), sin embargo en la mayoría (80%) de las especies usualmente es 64 (Moran, 2005). Sin embargo Bower (1926), menciona que el número de esporas por esporangio en la familia Hymenophyllaceae es extremadamente variable, reportando en sus resultados un rango de variación entre 32 y 421 esporas.
La monofilia de la familia Hymenophyllaceae no ha sido discutida y los análisis cladísticos están bien soportados (rbcL). Los caracteres sinapomórficos que agrupan a los taxones son dos, una lámina con una sola fila de células y esporas de color verde. Esta familia tiene cerca de 600 especies las mismas que podrían clasificarse tanto en 2 géneros como en 42 (Pacheco, L. & Riba, R., 1991). Aunque existe una evidente dicotomía basal para soportar a Trichomanes e Hymenophyllum como géneros, muchos pteridólogos, basándose en la diversidad morfológica de los taxones proponen la adición de más géneros tradicionalmente tratados como subgéneros (Moran, 2005).
Las mayoría de las especies de Hymenophyllaceae tienen una distribución Pantrópical, y en menor grado en regiones templadas, generalmente en bosques húmedos (Pacheco, L. & Riba, R., 1991). Principalmente son especies epífitas, pero también es posible encontrarlas como terrestres, epipétricas y a veces como hemiépifitas (Pacheco, 1995). Esta diversidad morfológica y ecológica, ha impulsado estudios generales sobre diversidad ecológica y tendencias de adaptación en los hábitos de los helecho tropicales (Dubuisson, 2003).
El propósito de este estudio fue hacer un relevamiento de las especies de Hymenophyllaceae más comunes en las estaciones biológicas de Las Cruces, Savegre, Cerro de la Muerte y la Selva para determinar si la variación en número de esporas tiene significancia taxonómica a nivel de los subgrupos de Trichomanes e Hymenophyllum.
Métodos
Sitios: La investigación se realizó en Costa Rica, en cuatro tipos de bosque: Tropical Pluvial Premontano ( Estación Biológica Las Cruces, 1180 m.s.n.m.), Montano húmedo (Albergue Montaña Savegre, 2100 m.s.n.m.), Páramo (Cerro de la Muerte, 3300 m.s.n.m.).y Bosque Tropical muy húmedo (Estación Biológica La Selva, 35-150 m.s.n.m.).
Trabajo de Campo: Los especimenes utilizados para el estudio fueron colectados en las caminatas por los principales senderos ecológicos en cada estación biológica visitada. El material vegetal utilizado para este estudio debía ser únicamente vivo y para ello se usó bolsas plásticas herméticas que mantuvieron los soros en un estado óptimo aproximadamente por 10 días.
Trabajo de Laboratorio: Para obtener el número de esporas, se realizó un conteo de las mismas en los esporangios que aún se encontraban cerrados y óptimos para el estudio. Se tomaron mínimo 2 individuos por especie, y los esporangios fueron extraídos de individuos y esporofitos diferentes tratando en lo posible que existan mínimo 2 réplicas por especie, en las mejores condiciones se lograron contar las esporas de 4 esporangios por especie. Los limitantes para estandarizar el tamaño de la muestra fueron las posibilidades de encontrar esporofitos fértiles con esporangios cerrados y el tiempo para realizar un conteo exacto. Para el proceso a nivel microscópico, se utilizaron técnicas de aislamiento para evitar que ingresen en el portaobjetos esporas liberadas de otros esporangios. El medio acuosos utilizado fue agua y en algunos casos especialmente con taxones del género Hymenophyllum, fue necesario usar azul de metileno para teñir esporas translúcidas. El poder de ampliación del microscopio que generalmente se usó fue el de 40 X y 100X, pero al observar esporas translúcidas o pequeñas fue necesario usar el lente de 400X.
Trabajo de herbario: Los ejemplares colectados fueron identificados y sus replicas fueron ingresadas en la base de datos de los herbarios del Instituto de Biodiversidad (INBio), Museo Nacional de Costa Rica (CR) y New York Botanical Garden.
Análisis y Mapeo filogenético: Los datos fueron registrados en una base de datos de Excel.
Para el mapeo cladístico del carácter número de esporas por esporangio, se utilizaron los cladogramas del género Trichomames (Dubuisson, en prensa 2005) e Hymenophyllum (Hennequin 2002).
Resultados
Se lograron encontrar 18 especies de Hymenophyllaceae. Dentro del género Trichomanes se encontraron 12 especies y en el género Hymenophyllum 6. La factibilidad para encontrar esporofitos maduros fue óptima a excepción de Trichomanes membranaceum.
El rango de variación del número de esporas por esporangio en las 17 especies observadas fue de 24 a 240 esporas. Los límites del conteo fueron representados por las especies Hymenophyllum hirsutum e H. tunbridgense
Quince especies presentaron variación en sus conteos, no obstante las especies constantes en su número de esporas fueron: Trichomanes elegans, T. collariatum, T. diaphanum e Hymenophyllum myriocarpum (Tabla 1).
Para la identificación morfológica de las especies se utilizaron claves botánicas de las referencias citadas. Todas las especies fueron identificadas sin mayor dificultad a excepción de Hymenophyllum fucoides e H. tunbridgense, especies cercanamente emparentadas (Mickel & Beitel 1998).
Tabla 1:
| Taxón | Subgéneros | Número de esporas por esporangio | Área de Estudio | Hábito | Sustrato |
| Hymenophyllum fucoides (Sw.) Sw. | Hymenophyllum | 52,91 | SA | Epífita | Angiospernma |
| Hymenophyllum fragile (Hedw) C.V. Morton | Spherocionium | 40,50 | LC | Epífita | Alsophila |
| Hymenophyllum hirsutum (L.) Sw. | Spherocionium | 24,56 | LC | Epífita | Alsophila |
| Hymenophyllum myriocarpum Hooker | Mecodium | 28,28 | SA, MU | Epifita, Epipétrica | Escallonia |
|
Hymenophyllum polyanthos
(Sw.) Sw var. polyanthos |
Mecodium | 66,76 | LC | Epífita | Alsophila |
| Hymenophyllum tunbridgense (L.) J. E. Smith | Hymenophyllum | 228, 240 | MU | Epífita | Escallonia |
| Trichomanes crispum Kunth in Humb. | Trichomanes | 51,52,52,56 | LC | Epífita | Alsophila |
| Trichomanes diaphanum Kunth in Humb. | Polyphlebium | 40,40 | LC | Epífita | Alsophila |
| Trichomanes elegans Rich. | Trichomanes | 57,57 | SE | Epipétrica |
Roca |
| Trichomanes godmanii Hooker | Dydimoglossum | 24,36 | SE | Epífita |
Welfia regia |
| Trichomanes kapplerianum Sturm in Mart. | Dydimoglossum | 41,44,51 | SE | Epífita | Rollinia pittieri |
| Trichomanes membranaceum L. | Dydimoglossum | 0 | SE | Epipétrica | Piedras grandes |
| Trichomanes polypodioides L. | Trichomanes | 28,28,56 | LC | Epífita | Alsophila |
| Tricomanes capillaceum L. | Callistopteris | 48,60 | LC, SA | Epífita | Alsophila |
| Tricomanes collariatum van den Bosch | Vandenboquia | 32,32 | SE | Hemiepífita; epipétrica | Pentachletra |
| Tricomanes krausii Hook.& Grev. | Dydimoglossum | 53,64 | LC | Epífita | Alsophila |
| Tricomanes radicans Sw. | Vandenboquia | 30,36,40 | SA, MU |
Epífita y Epipétrica |
Escallonia; en pequeñas cuevas. |
| Tricomanes reptans Sw. | Dydimoglossum | 44,48,51,64 | LC, SE | Epífita | Alsophila |
| LC:Estación Biológica Las Cruces; SA: Albergue Montaña Savegre; MU: Cerro de la Muerte; SE: Estación Biológica La Selva. |
Discusión
Los resultados obtenidos se relacionan con las observaciones hechas por Bower (1926) en cuanto a la variación en los rangos del número de esporas por esporangio dentro de los subgrupos de los géneros e incluso en la misma especie, siendo esto un carácter no común en los helechos.
El mapeo del carácter número de esporas por esporangio en el cladograma elaborado por Dubuisson (2005 en prensa) demostró que la variación de dicho carácter carece de significancia taxonómica tanto a nivel genérico como subgenérico (Figura 1); lo mismo sucedió con las pocas especies encontradas para el género Hymenophyllum al mapearlas en el árbol filogenético molecular de Hennequin (Figura 2). Sin embargo los resultados sirvieron inesperadamente para encontrar una marcada diferencia entre las especies emparentadas H. fucoides y H. tunbridgense
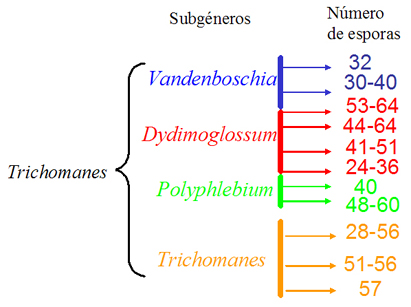
Figura 1: Mapeo
del número de esporas por esporangio en los sub-grupos del
género Trichomanes.
Cada rango pertenece a una especie, de las utilizadas para el
presente estudio.
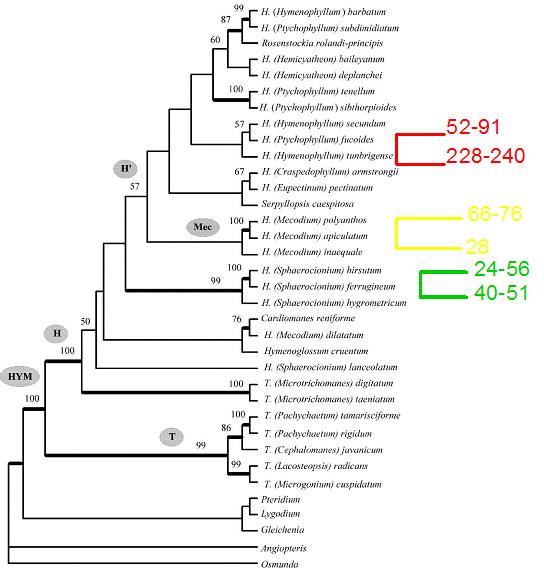
Figura 2: Mapeo
del número de esporas por esporangio sobre el cladograma del
género Hymenophyllum
(Hennequin et al. 2002).
Hymenophyllum fucoidesH. tunbridgense, "con frecuencia suelen ser confundidas" (Pacheco 1995) menciona que las distinciones morfológicas, al igual que los rangos geográficos, no se han resuelto en forma precisa.
La morfología comparativa de estas dos especies nos indica que la sinapomorfía que las emparentan, son últimos segmentos con márgenes aserrados o serulados; mientras que los caracteres que las dividen son: la orientación del soro respecto a la lámina, la longitud del pedúnculo y la anchura de los segmentos. Los estados de cada carácter podrían causar confusión e incertidumbre al ubicar a los especimenes en el taxón indicado. Tal es el caso del tamaño para la"anchura del segmento", carácter mencionado en la mayoría de las claves botánicas, en donde Hymenophyllum. tunbridgense es de 0.5 - 0.9 (1) mm. e H. fucoides es de 1-1.5 mm. (Mickel & Beitel 1998).
No obstante, si la comparación entre estas dos especies se realiza en función al carácter número de esporas por esporangio, la diferencia es marcada y amplia. H. fucoides presentó 91 esporas por esporangio como el valor más alto, mientras que H. tunbridgense presentó cuentas de 228 y 240. Además era notorio otras diferencias como el tamaño, y coloración de las esporas. Estos datos no han sido reportados en la literatura (Figura 3)
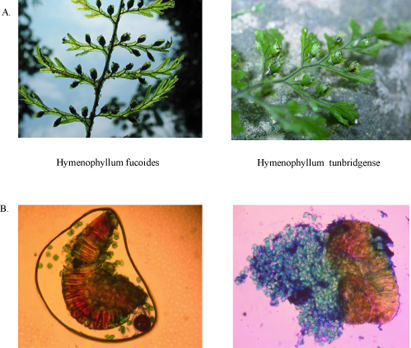
Es importante tener en cuenta que los resultados y discusiones versadas responden a un estudio exploratorio. Para futuras investigaciones que aborden este tema se recomienda: 1) ampliar el rango geográfico para descartar una significancia geográfica del carácter número de esporas por esporangio; 2) incrementar el tamaño de la muestra, con un mayor números de conteos de esporas por especie; 3) precisar datos en cuanto al color, forma y tamaño de las esporas para establecer una constancia en la diferencia entre H. fucoides e H. tunbridgense.
Referencias
Bower, F.O. 1926. The Ferns (Filicales). Vol. 2. Cambridge Univ. Press, United Kingdom. Pp: 243-244.
Dubuisson, J.Y. 2005. Árbol filogenético del género Trichomanes. Universidad de Paris, Francia, (datos en prensa).
Dubuisson J-Y.1; et al. 2003. Ecological Diversity and Adaptive Tendencies in the Tropical Fern Trichomanes L. (Hymenophyllaceae) with Special Reference to climbing and epiphytic habits. Botanical Journal of the Linnean Society. Vol.142. (1): 41-63(23).
Hennequin, S., A. Ebihara, M. Ito, K. Iwatsuki & J.-Y. Dubuisson. 2003. Molecular systematics of the fern genus Hymenophyllum s.l. (Hymenophyllaceae) based on chloroplastic coding and noncoding regions. Molecular Phylogenetic. 27: 283-301.
Lellinger, D. 1989. "The Ferns and Fern-alliens of Costa Rica, Panama, and the Chocó (Part 1: Psilotaceae through Dicksoniaceae)". Pteridología, 2A.: 185-228.
Mickel, J.T. & J. Beitel. 1988. Pteridophyta Flora of Oaxaca, Mexico. The New York Botanical Garden. Memoirs of the New York Botanical Garden, 46: 211-218.
Moran, R.C. 2004. A Natural History of FERNS. Pórtland-Cambridge. Timber Press. Inc. Portland, Oregon U.S.A. Pp: 15-42.
Moran, R.C. 2005. Géneros Neotropicales de Licofitos y Helechos. Una Guia para Estudiantes. Organización Para Estudios Tropicales, San José [manual fotocopiado para el curso OET 05-9].
Pacheco, L. & R. Riba. 1991. Hymenophyllaceae. Flora de Veracruz. Fascículo 63. Instituto de Ecología, A.C. Xalapa, Veracruz-Mexico. Editorial Futura S.A. Pág. 54.
Pacheco, L. 1995. Hymenophyllaceae. Pp. 62-83. En: R. C. Moran & R. Riba (eds). Flora de Mesoamérica, Vol. 1. Universidad Nacional Autónoma de México. Ciudad Universitaria, Mexico, D.F..