


 Email this Article
Email this Article Print
Print Download (PDF)
Download (PDF)
Resumen 
El proyecto se desarrolló en el Bosque de Mindo Lindo cerca
de Quito, el cual pertenece a la formación vegetal del bosque
húmedo premontano localizado entre la altitud de 1600 a 1800
m. La comunidad de hongos recolectada en"Mindo Lindo"
está constituida por Agaricales en un 62% y de
Aphyllophorales en un 37%. En la presente investigación sólo
se evaluó la diversidad y el análisis estructural de
los Aphyllophorales durante cinco meses en el año 2001 (Agosto
a Diciembre) en diez cuadrantes de -10 x 10 metros - dentro del
sotobosque. Se encontraron 132 individuos que representaban a 50
especies y 11 familias. La familia más importante fue
Polyporaceae con 12 especies, pero Corticiaceae, podría estar
en un futuro como la protagonista, si son revisadas las muestras por
los especialistas del grupo. La distribución y abundancia de
los hongos, depende de la "Cobertura Vegetal de Dosel",
como resultado la abundancia de los hongos es menor en los claros del
bosque. No se encontró una relación significativa entre
la precipitación y la abundancia de Aphyllophorales.
Abstract
The project was developed in Mindo Lindo, a premontane wet forest,
near Quito. Of the totalfungi collected in this area, 62% were
Agaricales and 37% were Aphyllophorales. The diversity and structural
analysis of Aphyllophorales was studied in ten plots -10 x 10
meters-, during five monthsfrom August through December in 2001-. A
total of 132 specimens belonging to 50 species in 11 families. The
most important family was Polyporaceae with 12 species, followed by
Corticiaceae, which could become increasingly important with better
identifications by specialists.
It was found that the distribution and abundance of fungi depended on
tree canopy cover, with less abundance in the forest gaps. A
significant relationship between rainfall and Aphyllophorales
abundance was not found.
Introducción
El detrimento del bosque nublado del noroccidente del Ecuador, lleva a la pérdida de todos los actores dentro de un ecosistema: plantas (productores), animales (consumidores), bacterias y hongos (descomponedores), si a esto se suman problemas ambientales globales como lluvia ácida y efecto invernadero, se puede establecer que ciertas comunidades de la naturaleza son más frágiles que otras. Un grupo muy sensible a la influencia ambiental, es el de los macrohongos. Si a esto se el descuido por parte de los ecuatorianos en desarrollar investigaciones en pro del conocimiento y conservación de los macrohongos, no sólo de este sector sino de todo el país, los hongos resultan ser los individuos más frágiles del ecosistema.
Estudios relativos a la diversidad de la micobiota, han sido desarrollados en Ecuador, en gran parte por extranjeros (Dumont et al. 1978), resultando algunos inclusive en el hallazgo de nuevas especies (Nishida 1989). De los macrohongos, los más renombrados son los Agaricales o Setas -forma de paraguas-. Sin embargo existe un grupo mucho más diverso en su forma, los Aphyllophorales, que dentro de las comunidades fúngicas son los que mantienen sus cuerpos fructíferos por más tiempo (Alexopolus et al. 1996), pero recibiendo muy poca atención por parte de los micólogos y botánicos (Zalamea 2002). Las investigaciones relativas a comunidades fúngicas de Aphyllophorales en los trópicos son pocas. El presente trabajo se fundamentó en las investigaciones de Lodge & Cantrell (1994), Lindblad (2001) y Zalamea (2000), en las que se analizan la influencia que la variación espacio temporal y el estado de sucesión de la vegetación tienen en la comunidad de macrohongos. La presente investigación trató de recoger, adaptar y unificar aspectos metodológicos de las investigaciones antes mencionadas, para determinar la diversidad y la estructura de la comunidad de los Aphyllophorales del Bosque Protector Mindo Lindo.
Materiales y Métodos
Mindo Lindo se encuentra en el kilómetro 79 de la carretera Calacalí - La Independencia, políticamente pertenece a la parroquia de Mindo del cantón Los Bancos en la provincia de Pichincha de Ecuador. Las coordenadas geográficas son 78°46´ W y 00°01´ S, con un rango altitudinal de 1600-1800 m. Es una reserva de propiedad de la Familia Peñafiel-Burnsehike, dedicada a la conservación, educación e investigación, este bosque está en recuperación desde hace aproximadamente 17 años.
Como unidad de muestreo, se definió a los basidiomas de Agaricales, los carpóforos de Aphyllophorales y a los Gasteromycetes. Estos hongos se colectaron dentro del sotobosque -hasta 2 m de alto- en diez cuadrantes de 10x10 m. durante los meses de Agosto a Diciembre del 2001, cubriendo parte de la temporada lluviosa y seca. Los sustratos donde fueron colectados los cuerpos fructíferos se marcaron con una cinta plástica naranja, para su revisión mensual.
De cada cuadrante, se graficó un "Perfil horizontal" de la comunidad de hongos, en hojas de papel milimetrado a escala 1:100. De esta manera cada individuo encontrado tuvo una ubicación en coordenadas cartesianas (x,y); obteniéndose datos de cada mes, que fueron sobrepuestos en AutoCAD 2002, logrando al final un estudio de perfiles horizontales de la estructura espacio-temporal de la comunidad fúngica.Para identificar la relación de la cobertura con la comunidad fúngica, se las ubicó según coordenadas (x, y), y se midió el tamaño de la ramificación -diámetro de copa- de los árboles que tenían un DAP > 10 cm. Este gráfico del perfil horizontal de la cobertura vegetal se sobrepuso al de los Aphyllophorales.
El tratamiento de las muestras de los hongos se la realizó en dos fases: colección e identificación
Las colectas de los cuerpos fructíferos de los Aphyllophorales y el resto de macrohongos se hizo en fundas de papel con los siguientes datos: color, tamaño, forma, sustrato, tipo de pudrición, datos ecológicos y fotografía a color del hongo. El secado de las muestras se realizó en el Herbario Nacional del Ecuador QCNE. El material traído a QCNE fue sometido a pruebas químicas con KOH, Floxina, Reactivo de Mezler y Azul de algodón y posteriormente se realizaron cortes para observar las características microscópicas y compararlas con los especímenes de la colección y las referencias bibliográficas. Las muestras se depositaron en el herbario QCNE, y con duplicados disponibles para intercambio.
Resultados y Discusión
Abundancia GeneralSe encontró 385 individuos de Agaricales, Aphyllophorales y Gasteromycetes, en los cinco meses, en los diez cuadrantes. De los grupos muestreados, los Agaricales presentaron el mayor procentaje de individuos - 62%- durante los 5 meses de análisis. Septiembre fue el mes más productivo con 113 individuos, y el acumulado del cuadrante No 5 con 62 individuos ([[Tabla 1]]).
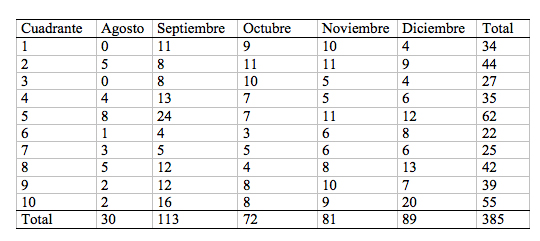
En la comunidad de hongos, la abundancia fluctúo mensualmente (Figura 1); F = 5,78 p = 0.075). Por esta razón durante el muestreo, se encontró por mes, un promedio de 77 (+ 30.37) individuos, que fluctuó con un máximo de 113 y un mínimo de 30 individuos por las características climáticas y la fenología de las especies. En cada cuadrante se estimó un promedio de 7.3 (+ 4.65) individuos y se estimó un promedio acumulado de 38.5 (+ 12.85) hongos por cuadrante. Se observó que variaba la abundancia de hongos de la micobiota de los cuadrantes, debido a la particularidad en cobertura y pendiente de cada uno.
Figura.1
Fluctuación de los individuos encontrados de Agaricales,
Aphyllophorales y Gasteromycetes
Figure 1. Individual fluctuation found at Agaricales, Aphyllophorales and Gasteromycetes.
Figure 1. Individual fluctuation found at Agaricales, Aphyllophorales and Gasteromycetes.
Abundancia de Aphyllophorales La abundancia durante los 5 meses, osciló entre 19 y 36 individuos, con un promedio de 26.2 (+ 8.02) individuos. Se considera que un solo individuo de los Aphyllophorales puede formar varios cuerpos fructíferos en el mismo tronco "sustrato" inclusive consume más de 2m2 o más del sustrato, eso sin contar con toda la degradación interna del tronco que realiza el micelio.
Las fluctuaciones en las poblaciones de Aphyllophorales, son menos notorias que la de los Agaricales, por las características de los cuerpos fructíferos de Aphyllophorales, de resistir más tiempo en el ambiente sin deteriorarse.
Análisis EstructuralLas poblaciones de hongos (Agaricales, Aphyllophorales y Gasteromycetes) dentro de cada cuadrante tienen una dispersión en agrupaciones o núcleos (S2 > X). Esto demuestra una interacción positiva de los individuos (Franco et al. 1985) que es influenciada por la acumulación de la materia orgánica, en descomposición - recurso indispensable para los hongos-. Aunque, no se logró establecer un patrón, fue notorio durante el trabajo de campo, que las agrupaciones de hongos estaban influenciados por la pendiente del terreno. Por estas variaciones topográficas el material orgánico -hojarasca- por acción de la lluvia y la erosión es arrastrado a sitios más planos, formando agregaciones de la comunidad de hongos.
Relación
con la Cobertura
En los cuadrantes se detectaron 330 individuos de Agaricales, Aphyllophorales y Gasteromycetes, de los cuales el 78.18% estaban dentro de la cobertura vegetal, ofrecida por los árboles (> a 10 cm. de DAP) y el 21.72% (+ 0.35) estaban fuera. Considerando los resultados de los análisis estadísticos, la cobertura vegetal influye de igual manera en ambos grupos de hongos, dando lugar a que la mayoría - 78.3 % - de hongos encontrados estén adaptados a la cantidad de luz y sombra del sotobosque. No hay una relación estadísticamente significativa de la abundancia de hongos y cobertura vegetal, obtenida de la formula CV = πab -en donde a y b son longitud de las ramas más grandes- (r2 = 0.232).
Los hongos no compiten por la luz, como las hierbas y otras plantas pioneras al aparecimiento de claros - gaps - (Smith 1995), pero si aumentan por el impacto producido en la caída de los árboles, ya que son las encargados de degradar la materia orgánica de los árboles caídos y regresar los nutrientes al ecosistema. A la variación espacio-temporal de la comunidad, se conoce con el nombre de parcelación (Krebs 1985). En el caso de la micobiota, este fenómeno se daría por el afloramiento de hongos y la dependencia al sustrato que degradan, y se ve influenciada por la topografía del terreno, factores fisonómicos - estructura, cobertura vegetal - del bosque o factores climáticos - lluvia, temperatura -. Por eso se consideró a la comunidad de hongos como una comunidad de no-equilibrio que cambia de acuerdo a la época climática "estaciones" y la dinámica del bosque. Este análisis es necesario manejarlo con cuidado, y se debe hacer hincapié en que la parcelación se basaría en el aparecimiento de nuevos individuos y no solo de fenología -aparecimiento de cuerpos fructíferos-.
La cobertura vegetal del dosel ayuda al desarrollo de los cuerpos fructíferos de los hongos, ya que la luminosidad del sol entra desde los lados (Smith 1995), y no desde arriba hacia el sotobosque, disminuyendo el impacto de la intensidad de los rayos solares. Gracias a esto se podría desarrollar técnicas para el manejo de los hongos dentro del bosque "productos forestales no maderables", y como consecuencia se evita la tala de bosque y se apoya a la conservación.
Diversidad
Durante los cinco meses de muestreo en los 1000 m2 se encontraron 142 individuos de Aphyllophorales, representados en 11 familias y 50 especies ([[Tabla 2]]), este número de especies es cercano a las 70 especies encontradas en un bosque alto andino de 30 años de recuperación (Zalamea 2000).
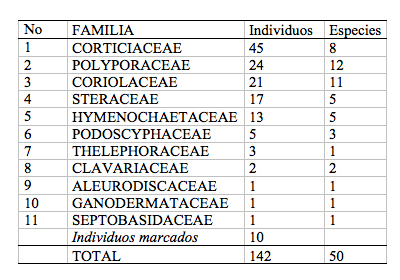
Las familias Polyporaceae y Corticiacae, fueron las más importantes dentro de la comunidad de Aphyllophorales del sotobosque de Mindo Lindo -considerando la diversidad relativa-. En la comunidad de Aphyllophorales las distribuciones de frecuencias son muy poco equitativas, con muchas especies "raras" - un solo individuo- y unas pocas especies "dominantes", esto concuerda con lo hallado en otras investigaciones (Zalamea 2000; Lindbland 2001). Es necesario aclarar que, para analizar la diversidad y abundancia de la comunidad fúngica se tomó en cuenta los cuerpos fructíferos de los Aphyllophorales. Se consideró además que las fluctuaciones durante los meses de muestreo están dadas por la fenología de la especie que esta influenciada por factores ambientales. (Dix & Webster 1985). Por lo que no se ha hecho mucho hincapié en los cambios de abundancia ni diversidad durante cada mes, porque en los sustratos pudieron encontrarse los hongos, pero no fueron tomados en cuenta en la encuesta respectiva por carecer de cuerpo fructífero.
Índice de Diversidad de Simpson
(IDS)
El Índice de Diversidad de Simpson (IDS) fue de 31.87 para 50 especies; esto indica que la comunidad de Aphyllophorales en este bosque se encuentra sobre el valor de medianamente diverso (Anexo, Tabla IV). Se excluyo a todo la familia Corticiaceae - por la dificultad que existió en su identificación y por ser la familia más abúndante-, y se obtuvo un IDS de 28.54 para 41 especies. Los Aphyllophorales se mantuvieron sobre el valor de medianamente diverso. Aunque se mantiene el nivel de la diversidad, excluyendo a Corticiaceae, esta familia no deja de ser la más importante dentro de la micobiota de los bosques tropicales (Lindbland 2001). Basado en los resultados obtenidos se afirma que la comunidad de Aphyllophorales del Bosque protector Mindo Lindo durante los meses de Agosto a Diciembre del 2001 se encuentra sobre el valor de medianamente diverso, pero el número de especies podría aumentar al perfeccionar las determinaciones taxonómicas inclusive, con la posibilidad de encontrarse especies nuevas.
Sustratos y Pudrición de Madera
No hay una relación estadísticamente significativa entre el Volumen de madera (m3) y la abundancia de hongos (r2=0.166). Esto se debe a que los hongos encontrados no solo degradan madera sino también hojarasca y ramas pequeñas (> 2.5 cm. de diámetro) y por supuesto troncos (> 10 cm. de diámetro). La mayoría de hongos que se encontró estaban sobre ramas "Humícolas" (> 2.5 cm. de diámetro), muy pocos en troncos "Lignícolas" (> 10 cm. de DAP). Además se halló tres individuos atacando las raíces y la parte inferior del tronco que soportaba un "matapalo" de Clusia (Clusiaceae), llegando inclusive uno de ellos a caer durante el trabajo de campo por la degradación producida por el hongo en las raíces del árbol. El tipo de pudrición de la mayoría de hongos encontrados fue la pudrición blanca, que degrada celulosas como lignina y que se observa como fibras blancuzcas. (Dix & Webster 1995; Zalamea 2002).
Para mantener constantes los ciclos de los nutrientes, los hongos tienen que ir degradando los sustratos y hasta que no terminen de degradar, todo permanecerá en este sustrato, en ocasiones con un cuerpo fructífero visible y en otras no (fenología); para luego migrar a otro sustrato por medio de las esporas. Por estas razones, fueron notorias las oscilaciones de la abundancia, por el aparecimiento de nuevos individuos durante el muestreo. Es necesario tomar en cuenta que la diversidad y análisis estructural de la micobiota que se presenta, podría cambiar después de unos años, según la dinámica del bosque.
Los resultados demuestran la necesidad de investigaciones más detalladas sobre los macrohongos del Ecuador, tanto en aspectos como Ecología, Fenología y Composición de la micobiota y taxonomía, se sustentan con los resultados obtenidos, "nada aún esta dicho". Es importante tener más información sobre los descomponedores de materia orgánica de nuestros bosques, de esta manera se entenderá mucho mejor a los ecosistemas.
Agradecimientos
A David Neill, Mercedes Asanza, Tatiana San Juan, Ximena Aguirre y a todo el personal del Herbario Nacional de Ecuador (QCNE), por su apoyo, y por condescenderme la Beca "Elizabeth Neill", que permitió el desarrollo de este proyecto. A los especialistas Ewald Langer, Roman Kuetter, Fernando Fernández, Mitzi Campo Agüero, Thomas Læssøe, por el material para el desarrollo del trabajo de laboratorio, literatura, apoyo, sugerencias, y revisión de los especimenes, respectivamente.
Referencias
Alexopolus, C.; C. Mims & M. Blacwell. 1996. Introductory Mycology. John Wiley & Sons. Inc.
Dix, N. & J. Webster. 1995 Fungal Ecology, Chapman & Hall, Estados Unidos
Donk, M. 1971. Multiple Convergence in the Polyporaceae. Pp. 393-422. En: R. Petersen. Evolution in the Higer Basidiomycetes. University of Tennessee Press, Knoxville, TN.
Dumon, K., Buritica, P. & E. Forero. 1978. Los Hongos de Colombia I, Introducción Caldasia Vol. XII, No. 57.
Harmon, M.; E. Whigham; D. Sextio & I. Olmsted. 1995. Decomposition and mass of woody detritus in the dry tropical forest of the northeastern Yucatan Peninsul, Mexico, Biotropica 2: 305-316
Hedger, J.; D. Lodge; G. Dickson; H. Gitay; T. Laessoe. & R. Watling. 1995. To The Bms Expedition Cuyabeno Ecuador: And Introduction Mycologist vol.9, Part 4.
Lindbland, I. 2001.Diversity of poroid and some croticioid wood-inhabiting fungi along the rainfall gradient in tropical forests, Costa Rica. Journal of Tropical Ecology 17: 353-369. University Press
Lodge, J. & S. Cantrell 1995. Fungal communities in wet tropical forests: variation in time and space. Can. J. Bot. 73 (Suppl.1): 1391-1398
Krebs, C. 1985. Ecología, Análisis Experimental de la Distribución y Abundancia. Ediciones Pirámide S.A. Madrid.
Nishida, F. 1989. Review of Mycological Studies in the Neotropics. En: Campbell, D. & D. Hammons. (eds.) Floristic Inventory of Tropical Countries. The Status of Plant Systematics, Collections and Vegetation; plus. Recomendations for the future. New York Botanical Garden 1989.
Zalamea, M. 2000 Estudio de la comunidad de hongos lignícolas en dos estadios sucesionales de un bosque altoandino. Trabajo de grado. Biología. Universidad Nacional de Colombia
Zalamea, M. 2002. Guía para recolectar hongos, macromicetos del orden Aphyllophorales. En Perez (Ed.) Arbelaezia No. 13, Febrero 2002, Colombia