


 Email this Article
Email this Article Print
Print Download (PDF)
Download (PDF)
Resumen 
El manejo de los bosques nativos y en especial de los bosques
tropicales de tierras bajas es una necesidad urgente en Ecuador. Tal
propósito es contrarestado por limitantes sociales, económicas
y hasta técnicas. Este documento se basa en el análisis
de aproximadamente 40 000 árboles medidos en 10 500 ha en el
noroccidente de Ecuador. A partir de esos datos se analizó la
estructura del bosque y se agruparon las especies por gremios
ecológicos. Como resultado de los inventarios, se registraron
265 especies mayores a 10 cm de DAP; de estas, 74 se identificaron
hasta especie, 79 hasta género, y 112 se reconocieron solo por
sus acepciones comunes. De las especies identificadas hasta especie y
género (153), tres fueron clasificadas como heliófitas
efímeras, 31 como heliófitas durables, 94 como
esciófitas parciales y tres como esciófitas totales.
Referente a la distribución por clases diamétricas, las
especies esciófitas presentaron una típica J-invertida
con una alta concentración de individuos en clase 10, mientras
que las heliófitas mostraron una distribución casi
horizontal. Esto se explica porque las esciófitas tienen la
capacidad de regenerarse bajo la sombra, mientras las heliófitas
aprovechan las bondades de los claros para crecer rápidamente.
Los resultados también mostraron que el 40 % de las especies
superan los 60 cm de DAP, límite oficial de corta de la
mayoría de especies. Al relacionar los usos actuales, se
observó que las especies usadas para desenrollo son en su
mayoría esciófitas con una buena distribución
diamétrica. En cambio, las heliófitas durables de
rápido crecimiento y consideradas valiosas, tienen una
distribución horizontal.
Abstract
Natural forest management (NFM), particularly of lowland tropical
forest, is urgently needed in Ecuador. However, there are social,
economic and technical limitations. This document is based on the
analysis of inventory data of approximately 40 000 trees from 10 500
hectares in the North-west of Ecuador, an area which provides at
least 60% of the timber used by the country. Using these data forest
structure was analysed and species grouped to ecological
characteristics. In the inventories 265 species were registered above
10 cm diameter breast height (DBH); of these, 74 were identified to
species level, 79 to genus and 112 by local names. Of the 153 species
identified to species or genus level, three were classed pioneers, 31
as long-lived light demanders, 94 as partial shade-bearers and three
as total shade-bearers. The shade-bearing species showed a typical
negative exponential diameter class distribution while that of the
light-demanders was almost horizontal. This is explained as the shade
tolerant species have the capacity to regenerate in shade, while the
light demanders do so in the gaps, without being able to regenerate
in their own shade. The results also show that 40% of species exceed
60cm DBH, the official minimum cutting diameter of the majority. This
information is key to determining the minimum cutting diameters when
growth data is lacking, as in the case of Ecuador. Actual timber use
shows that most peeler species are shade-bearing with a good diameter
class distribution. On the other hand the long-lived light demanders,
typically relatively fast growing valuable timbers, show a flat
diameter class distribution making them more vulnerable. This
characteristic is also shown by some shade bearers like chanul, the
most valuable species in the area, which has a flat diameter class
distribution.
Introducción
A partir de promulgación de las Normas para el Manejo Forestal Sustentable de los bosques tropicales húmedos del Ecuador (MAE 2000), ha surgido un verdadero impulso al manejo de los bosques nativos. Pero, a más de los aspectos políticos y legales que son de ámbito coyuntural, el manejo forestal requiere superar una serie de limitantes de orden social, económico y técnico. Entre los limitantes técnicos, un aspecto de enorme importancia es ¿cómo simplificar la silvicultura de bosques ricos en especies cuando en la práctica se aprovechan pocas especies, o son pocas las que tienen mercado?
Algunos forestales y ecólogos forestales han propuesto que para manejar los bosques, debe considerarse la agrupación de las especies de acuerdo a características ecológicas para así simplificar su manejo.
Los bosques nativos ecuatorianos están entre los más ricos y diversos del mundo. En este marco, los bosques tropicales húmedos, los más extensos del país, también difieren significativamente en composición florística y riqueza de madera (ver Palacios & Jaramillo 2001).
Como un aporte al manejo de este tipo bosques, se hizo el análisis de inventarios forestales de más de 10500 hectáreas de bosques en el noroccidente del Ecuador. Con ese fin, se agruparon las especies bajo diferentes criterios.
Materiales y Métodos
Área de estudio
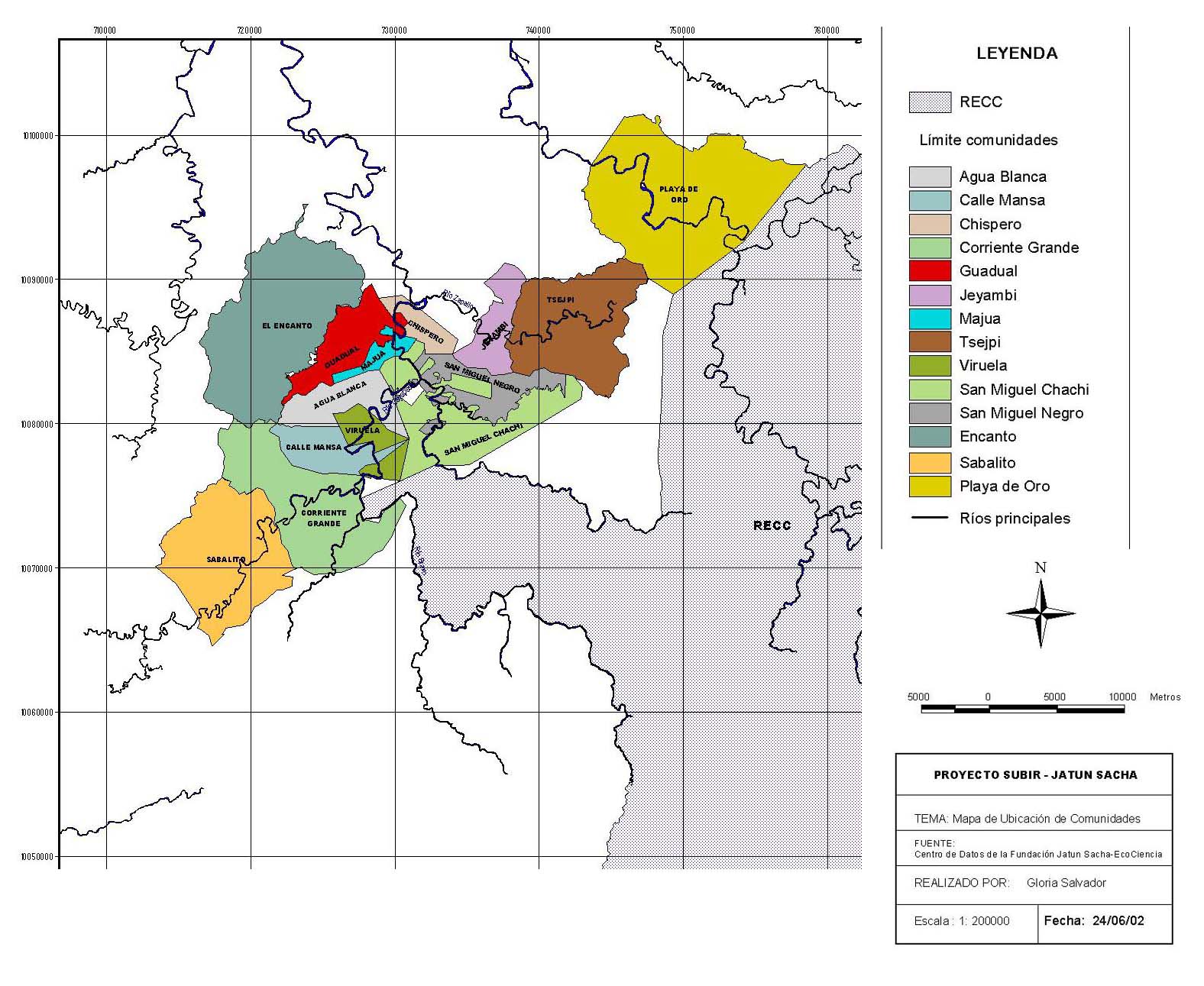
Figura 1. Comunidades del Río Cayapas donde se
realizaron inventarios forestales.
Figure
1. Communities of thye study area along Rio Cayapas.
El área de estudio (ver (Figura 1) se ubica en la provincia de Esmeraldas, noroccidente de Ecuador, río Cayapas. La zona es parte del bosque muy húmedo tropical (Cañadas 1981) o un bosque siempre verde de tierras bajas (Cerón et al. 1999). Concentra más de 110 especies arbóreas por hectárea donde, precisamente, la mayor riqueza florística no se concentra en este hábito, sino en las especies epífitas, hemiepífitas y plantas del sotobosque (Palacios & Jaramillo 2001). Los suelos son ácidos y de baja fertilidad. Las condiciones climáticas están caracterizadas por temperaturas sobre los 24 ºC y elevadas precipitaciones; aunque, en los últimos años se observó una fuerte disminución de lluvias entre agosto y diciembre.
La cuenca del río Cayapas es particularmente importante por la gran cantidad de madera que oferta. Hasta 1993, el 80 % de la madera que se consumía en el Ecuador salía de Esmeraldas (ITTO/INEFAN 1993). En los últimos años la tendencia ha cambiado ligeramente, con la ampliación de la frontera de extracción forestal en todo el país.
Los datos para el presente documento provienen de siete comunidades de la parte media del río Cayapas, asentadas entre 50 y 250 m sobre el nivel del mar.
Toma de datos
Los inventarios forestales se realizaron para levantar información de las características del bosque y del terreno durante el proyecto SUBIR (Sustainable Use of Biological Resources). Se aplicaron muestreos sistemáticos en 9 000 ha de bosque primario primarios no intervenidos o poco intervenidos a intensidades entre 0.6 hasta 7.0 % ([[Tabla 1]]). En todos los inventarios se incluyeron las palmas, y los errores de muestreo estuvieron por debajo del 20 %. Se midieron alrededor de 40 000 árboles.
En Guadual (Jaramillo & Palacios 1998), Chispero (Tipaz & Zuleta 1998), Calle Mansa (Prado 1999) y Jeyambi (Pozo & Garrido 1998) se utilizaron parcelas de 20 x 500 m y se midieron los árboles mayores a 10 cm de dap; en Tsejpi (Obando & Tipaz 1998) se practicaron dos inventarios: el primero con parcelas de 20 x 500 m y el segundo con parcelas circulares de 500 m² y se midieron los árboles mayores a 10 cm de dap (Obando 1999). Y finalmente, en El Encanto (Zuleta & Palacios 2001) y Corriente Grande (Garrido 2002) se usaron fajas de 10 m de ancho por un largo indeterminado con estimador de razón (ver Jolitz & Palacios 2000). En cada faja se midieron los individuos sobre 60 cm de dap; dentro de cada faja se establecieron parcelas de 10 x 400 m en donde se midieron todos los individuos mayores a 30 cm de dap; mientras que, en una subparcela de 10 x 100 m ubicada en el extremo de cada parcela se midieron los individuos mayores a 10 cm de dap. Para todos los individuos se tomó la altura comercial.
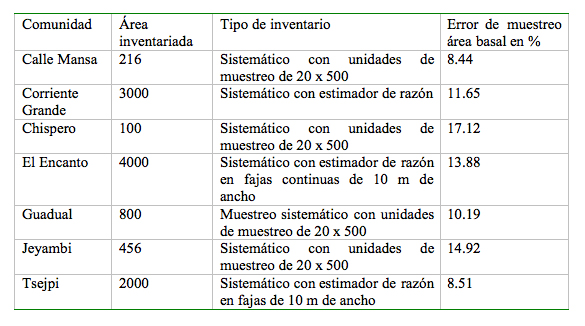
Tabla 1. Comunidades, áreas
tipo de inventario y error de muestreo empleadas por el Proyecto
SUBIR.
Table 1. Communities, tipical investigation areas and sample error applies for Project SUBIR.
Table 1. Communities, tipical investigation areas and sample error applies for Project SUBIR.
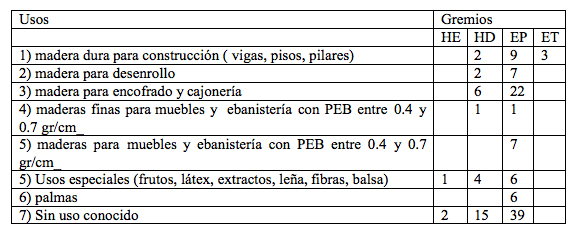
Tabla 2. Número de especies
agrupadas por gremios ecológicos y por tipo de uso.
Table 2. Species numbers by ecological groups and useage type.
Table 2. Species numbers by ecological groups and useage type.
Criterios para la agrupación de
especies
Las especies se agruparon bajo tres variables: gremio ecológico, distribución diamétrica y uso. Para la agrupación en gremios se usó la propuesta de Finegan & Delgado (1997) empleada por Louman et al., (2001). Tal agrupación se aplicó a todas las especies identificadas hasta el nivel de especie y género. Las especies identificadas únicamente por las acepciones comunes no se clasificaron.
Para conocer los aspectos de la estructura horizontal, todos los árboles se agruparon en clases diamétricas de tamaño 10 (clase 10: 10.0-19.9 cm y así sucesivamente) en función de los gremios. Y con respecto a la agrupación por usos, se consideraron siete clases aplicadas a las especies identificadas a nivel de especie y género, a saber:
1) Maderas para construcción (vigas, pisos, tumbados, etc.) generalmente, con un peso específico básico (PEB) por encima de 0.40 gr/cm³
2) Maderas para desenrollo que incluyen especies de madera suave y semiduras, usualmente con daps por encima de 55 cm.
3) Maderas para encofrado y cajonería. Maderas suaves y semiduras con daps por encima de 40 cm.
4) Maderas finas para muebles y ebanistería, con PEB entre 0.40 y 0.75 gr/cm³.
5) Maderas para muebles y ebanistería con PEB por debajo de 0.40 y 0.75 gr/cm³.
6) Usos especiales. Dentro de este grupo se incluyeron especies que producen frutos comestibles (Inga spp.), fibras (Poulsenia armata), látex (Castilla elastica), leña (Huberodendron patinoi) y piezas para balsas (Ochroma pyramidale, Trichospermum galleottii) de transporte de madera aserrada o rolliza.
7) Palmas. En este grupo se incluyeron todas las palmas de tamaño arbóreo, usadas como varengas para techos, parquet, muebles, construcción en general y artesanías.
0) Maderas sin uso. En este grupo se incluyen todas las especies sin uso conocido, conformado principalmente por especies del subdosel, o por especies que debido a su baja abundancia no son conocidas, y por tanto, tampoco se usan.
Resultados
[[Composición florística y gremios ecológicos]]
De 265 especies registradas en los inventarios, 74 fueron identificadas hasta especie, 79 hasta género, mientras que 112 (42 %) solo se reconocieron por sus acepciones comunes o fueron desconocidas ([[Tabla 3]]). De las especies identificadas a nivel de especie y a nivel de género (153), tres especies fueron clasificadas como heliófitas efímeras, 31 como heliófitas durables, 94 como esciófitas parciales y tres como esciófitas totales. Estos datos contrastan con los reportados por Hartshorn (1980) y Werner (1986) citados por Finegan (1993), y Finegan y Sabogal (1988), quienes advierten un mayor número de especies heliófitas que esciófitas, con una mayor abundancia de individuos de las segundas, en bosques tropicales húmedos de Costa Rica (Finegan 1993). El mayor número de especies esciófitas puede explicarse por la facilidad que tienen para regenerarse en cualquier fase del ciclo de regeneración (Finegan & Delgado 1996), incluyendo condiciones de sombra. En nuestro estudio, el número de esciófitas también es mayor por la inclusión de muchas especies que crecen y permanecen en el subdosel y son consideradas esciófitas.
Un aspecto que incide en la actual estructura y composición de los bosques analizados, es la historia de los asentamientos humanos. La población Chachi y Negra está en la zona del Cayapas menos de 500 años (Montenegro & Bonifaz 1996), mientras que los asentamientos más antiguos sólo se localizaron en la costa misma. Así, estos bosques parecen ser muy maduros, y no sujetos a las intervenciones antrópicas recientes, característica de muchos bosques tropicales en los últimos cientos de años (Finegan 1993).
Pese al mayor número de especies esciófitas en los bosques del noroccidente de Ecuador, todavía hay un importante número de especies heliófitas durables que crecen rápido, tienen madera de buena calidad, lo que las hace muy favorables al manejo.
Distribución por clases
diamétricas
La distribución por clases diamétricas a nivel de gremios ecológicos aparece en la Figura 2. Se ve claramente que la tendencia por grupos de especies varía considerablemente. Las especies esciófitas presentan una típica J-invertida con una alta concentración de individuos en las clases diamétricas inferiores (138 individuos por hectárea en la clase 10 (10-19.9 cm)).Tales individuos pertenecen a especies esciófitas que superan los 60 cm de dap en edad adulta, pero también a especies del subdosel que no alcanzan diámetros superiores, y que son tolerantes a la sombra. Dentro de este grupo están las palmas, las cuales son muy abundantes (Wettinia quinaria e Iriartea deltoidea con 40 y 23 individuos /ha, respectivamente) en el noroccidente de Ecuador aunque no sobrepasan los 30 cm de dap. El gran número de individuos esciófitos en las clases por debajo de 30 y la reducción brusca a partir de este límite, indicaría una mortalidad alta en esas clases (Finegan & Delgado 1996). Por otro lado, la presencia de individuos esciófitos (que alcanzan grandes tamaños) en las clases diamétricas bajas, no siempre indica juventud; puede suceder que esos árboles sean ya adultos, aunque permanezcan en el subdosel.
Las heliófitas efímeras y esciófitas totales muestran una distribución muy similar. Parece ser que para las primeras, mantienen poblaciones más o menos estables cuando superan los 10 cm, pues sobre este límite los árboles se vuelven vigorosos (Finegan & Delgado 1996) y todavía siguen disfrutando de las ventajas de crecer el un claro. La distribución horizontal de ciertas especies esciófitas como el chanul, puede deberse a una escasa regeneración natural (Terán, 2002) o a tasas altas de mortalidad en las clases diamétricas bajas. Este podría ser el caso del guadaripo (Nectandra guadaripo). Este tipo de distribución diamétrica complica las opciones de manejo; por un lado, son especies altamente demandadas por su madera dura, y por otro, existe la probabilidad de no reponer los árboles que se cosechan por falta de regeneración.
Con respecto a las heliófitas durables, 21 especies superan los 60 cm de dap, pero de éstas, cinco (Cedrela odorata, Casearia arborea, Vochysia macrophylla, Hyeronima alchorneoides y Apeiba membranacea) son las especies con mayor importancia comercial con 2.1 árboles por ha mayores a 60 cm de dap. La distribución diamétrica de este gremio es dramáticamente diferente al de las esciófitas. El número de individuos en la clase 10 es de 12.5 árboles por hectárea, disminuye a 9.8 en clase 20 y luego se mantiene más o menos horizontal.
Otro aspecto analizado fue el diámetro máximo de las especies. Del total de especies (265), el 75 % superan los 40 cm de dap, un 40 % supera los 60 cm y un 22 % no pasa la clase de 10 cm. El 25 % del total de las especies que no alcanzan los 40 cm de dap, son especies típicamente del subdosel y que en ámbito comercial tendrían poco uso. El límite de 60 cm es el diámetro mínimo de corta (dmc) oficial para de la mayoría de especies (MAE, 2000). Un ajuste del dmc oficial para los bosques del noroccidente es posible con los datos de este estudio.
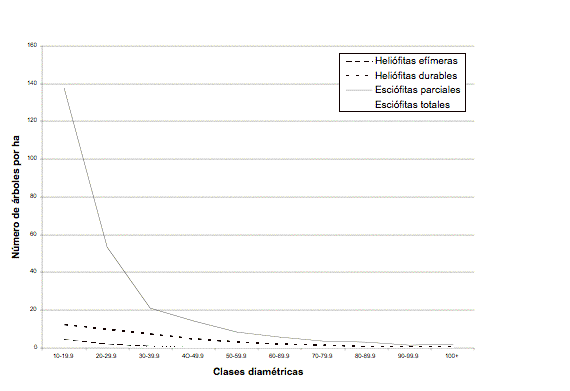
Figura 2. Distribución diamétrica por
gremios de especies, noroccidente de Ecuador.
Figure
2. Diameter distribution of different species assemblages in
Northwestern Ecuador.
Relación entre los gremios ecológicos
y los usos comerciales
La relación entre los gremios ecológicos y los usos de las especies aparece en las [[Figuras 3, 4, 5 y 6]]. Las esciófitas totales se usan principalmente para construcción, lo cual complica la situación de esas especies por la alta demanda que tienen y por los problemas silvícolas. El grupo de las esciófitas parciales con siete usos, presenta mejores perspectivas, en especial de las especies dedicadas a desenrollo y con una distribución diamétrica regular. Para este grupo ecológico, se promedió más de 7.5 árboles/ha por encima de 60 cm de dap y 36 árboles por debajo de ese límite. En este grupo de especies se incluyen sande (Brosimum utile), chalviande (Virola reidii y V. dixonii), cuángare (Otoba gordoniifolia y O. gracilipes) y pulgande (Dacryodes sp.). El sande con 3.5 árboles ≥ 60 cm de DAP por hectárea es la especie más abundante de este grupo, y en general de todas las especies arbóreas comerciales del noroccidente. Le sigue el guadaripo (Nectandra guadaripo) con 1.6 árboles.
Las especies heliófitas durables, entre las cuales constan el cedro (Cedrela odorata), mascarey (Hyeronima alchorneoides), carrá (Huberodendron patinoi) y azafrán (Zanthoxylum riedelianum) son poco abundantes en el bosque primario. La especie más abundante en este grupo es peine de mono (Apeiba mebranacea), una especie de madera muy suave y usada como decorativa, con 1.3 árboles/ha. La siguiente especie en importancia es mascarey con 06 árboles/ha. La especie con la madera más fina en este grupo, el cedro, no se encuentra por encima de los 60 cm de dap. Y esta es una característica general de los bosques de bajura del noroccidente del Ecuador: ricos en especies, pero pobres en especies altamente comerciales (Palacios & Jaramillo 2001).
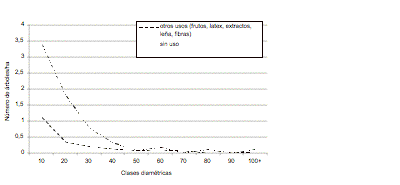
Figura 3. Distribución
diamétrica de las heliófitas efímeras por tipo
de uso
Figure 3. Diameter distribution of ephemerous heliophytes according to use.
Figure 3. Diameter distribution of ephemerous heliophytes according to use.
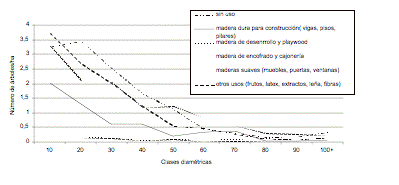
Figura 4. Distribución
diamétrica de las heliófitas durables por tipo de uso.
Figure 4. Diameter distribution of heliophytes according to use.
Figure 4. Diameter distribution of heliophytes according to use.

Figura 5. Distribución
diamétrica de las esciófitas parciales por tipo de uso.
Figure 5. Diameter distribution of partial esciophytes according to use.
Figure 5. Diameter distribution of partial esciophytes according to use.
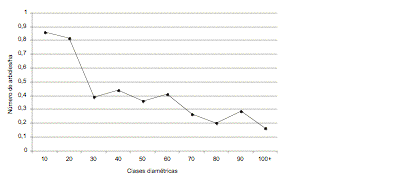
Figura 6. Distribución
diamétrica de las esciófitas totales por tipo de uso.
Figure 6. Diameter distribution of complete esciophytes according to use.
Figure 6. Diameter distribution of complete esciophytes according to use.
Discusión
Como se ha mencionado, los bosques del noroccidente del Ecuador son altamente diversos y como tales, requieren de criterios rigurosos de manejo para mantener al máximo sus características. También es claro, que cualquier intervención forestal implicará cambios en la composición florística y en la estructura del bosque, así como en las características físicas del área. No obstante, es también concluyente que cualquier forma de manejo forestal es mejor que una pastura o un monocultivo de palma africana.
La simplificación de los bosques tropicales húmedos para posibilitar su manejo ha sido un tema ampliamente tratado (ver Lamprecht 1990). Para llegar a tal simplificación se han practicado una serie de tratamientos silvícolas. Dejando de lado a la cosecha, dos de los tratamientos más sobresalientes son el refinamiento y la liberación (ver Hutchinson 1993, Quirós 2001a). El primero de estos, contribuye a la eliminación de individuos por razón exclusivamente de su especie (Hutchinson 1993), lo que constituye la práctica de una silvicultura negativa; mientras que la liberación quita del bosque a los individuos que únicamente compiten con aquellos llamados "deseables".
Una forma de mejorar la calidad del bosque es aplicar tratamientos silvícolas en función de los gremios (ver Louman, et al. 2001). Si la teoría es válida, cada gremio tendría un comportamiento frente a la luz, crecimiento y características de la madera distintos. Así los tratamientos se orientarían a favorecer a un conjunto de individuos de un gremio de especies en particular. Aunque, eventualmente, se podría trabajar con más de un gremio aplicando un ciclo de corta para un gremio principal y ciclos de corta múltiplos o submúltiplos del primero para el resto de gremios. Por ejemplo ciclos de corta de 15 años para las heliófitas durables y 30 años para las esciófitas.
La agrupación en gremios ecológicos también permite la simulación del crecimiento del bosque. Si se asume que las especies de un gremio crecen de manera similar, con información disponible de especies representativas de un gremio, se puede hacer proyecciones cercanas a la realidad, sobre todo, cuando no se dispone de datos de crecimiento de largo plazo.
En resumen, bosques primarios discetáneos dominados por especies esciófitas como aquellos del noroccidente del Ecuador, deben ser sometidos a sistemas policíclicos. La perversa conclusión que gran parte de los bosques del norte de Esmeraldas son secundarios no tiene ningún fundamento, y a muchas luces se demuestra que no lo son. Y aún, si fueron intervenidos hace 25 o más años, la estructura y composición florística actual muestran que deben ser sometidos a sistemas policíclicos de manejo. En Centroamérica, en áreas afectadas por los grandes huracanes, también se ha optado por estos sistemas (Quirós 2001b). Estos sistemas mantienen la estructura discetánea del bosque (Quirós 2001b), característica muy visible cuando se mira la distribución diamétrica.
Agradecimientos
Todos los datos usados para este estudio fueron generados bajo el Proyecto CARE-SUBIR dentro del Componente Mejor Uso de la Tierra ejecutado por la Fundación Jatun Sacha, mismo que fue financiado por USAID. Agradecimientos también para el Proyecto Conservación de Áreas Indígenas Manejadas (CAIMAN) y a David Thomas por la revisión del documento.
Referencias
Cañadas, L. 1983. El Mapa Bioclimático del Ecuador. Banco Central del Ecuador.
Cerón, C.; W. Palacios; R. Sierra & R. Valencia. 1999. Las Formaciones Vegetales de la Costa del Ecuador. En Sierra, R. (ed.). Propuesta Preliminar de un Sistema de Vegetación para el Ecuador Continental. Proyecto INEFAN/GEF-BIRF y Ecociencia. Quito, Ecuador.
Finegan, B. & Sabogal, C. 1988. El desarrollo de sistemas de producción sostenible en los bosques húmedos de bajura. Un estudio de caso en Costa Rica. Parte 2. El Chasqui 18: 16-24.
Finegan, B.1996. Bases ecológicas para la silvicultura. CATIE.
Finegan, B & D. Delgado. 1996. Bases Ecológicas para el Manejo de Bosques Tropicales: Los ambientes forestales tropicales y el ajuste de las especies vegetales. CATIE.
Finegan, B & D. Delgado. 1997. Bases Ecológicas para el Manejo de Bosques Tropicales. CATIE.
Garrido, N. 2002. Plan de Manejo Integral de Corriente Grande. Proyecto SUBIR. Jatun Sacha. Fundación Jatun Sacha/Proyecto SUBIR. Quito, Ecuador
Hutchinson, I. 1993. Puntos de Partida y Muestreo Diagnóstico par la Silvicultura de Bosques Naturales del Trópico Húmedo. CATIE. Turrialba, Costa Rica.
Internacional Tropical Timber Organization/Instituto Ecuatoriano Forestal. 1993. Estrategias para la industria sostenida de la madera en el Ecuador. Proyecto PD 137/91. INEFAN. Quito.
Jaramillo, N. & W. Palacios. 1998. Plan de Manejo Forestal Comunitario de Guadual.
Proyecto SUBIR. Jatun Sacha. Fundación Jatun Sacha/Proyecto SUBIR. Quito, Ecuador
Jolitz, T. & W. Palacios. 2000. Manual para Inventarios Forestales. Fundación Jatun Sacha/Proyecto SUBIR. Quito, Ecuador.
Lamprecht, H. 1990. Silvicultura en los trópicos. Cooperación Técnica-República Federal de Alemania (GTZ). Eschborn.
Louman, B.; J. Valerio & W. Jiménez. 2001. Bases ecológicas. En Louman, B., D. Quirós & M. Nilsson (eds.). Silvicultura de bosques latifoliados húmedos con énfasis en América Central. CATIE. Turrialba, Costa Rica.
Ministerio del Ambiente del Ecuador. 2000. Normas para el manejo forestal sustentable para aprovechamiento de madera en bosque húmedo. Quito, Ecuador.
Montenegro, F. & M. Bonifaz, M. 1996. Plan de manejo Forestal del Centro Chachi Pichi yacu Grande. Fundación Juan Manuel Durini. Quito, Ecuador.
Obando, T. & Tipaz, G. 1998. Plan de Manejo Forestal Comunitario del Centro Chachi Tsejpi. Jatun Sacha/Proyecto SUBIR. Quito, Ecuador.
Obando, T. 1999. Plan de Manejo Integral Comunitario del Centro Chachi Tsejpi. Jatun Sacha/Proyecto SUBIR. Quito, Ecuador
Palacios, W. & N. Jaramillo. 2001. Riqueza florística y forestal de los bosques tropicales húmedos del Ecuador e implicaciones para su manejo. Turrialba, 36: 46-50.
Pozo, N. & N. Garrido. 1998. Plan de Manejo Forestal Comunitario de Jeyambi. Jatun Sacha/Proyecto SUBIR. Quito, Ecuador.
Prado, L. 1999. Plan de Manejo Integral Comunitario del Centro Chachi Calle Mansa. Jatun Sacha/Proyecto SUBIR. Quito, Ecuador.
Quirós, D. 2001. Tratamientos silviculturales. En Louman, B., D. Quirós & M. Nilsson. Silvicultura de bosques latifoliados húmedos con énfasis en América Central. CATIE. Turrialba, Costa Rica.
Quirós, D. 2001b. Sistemas silviculturales. En Louman, B., D. Quirós & M. Nilsson. Silvicultura de bosques latifoliados húmedos con énfasis en América Central. CATIE. Turrialba, Costa Rica.
Tipaz, G. & J. Zuleta 1998. Plan de Manejo Forestal Comunitario de la Comunidad Chispero. Jatun Sacha/Proyecto SUBIR. Quito, Ecuador.
Zuleta, J. & W. Palacios. 2001. Plan de Manejo Forestal Comunitario de El Encanto. Jatun Sacha/Proyecto SUBIR. Quito, Ecuador.
Tabla 3. Lista de especies registradas en los inventarios forestales del Río Cayapas, Noroccidente del Ecuador.
Table 3. List of species encounteres in the forest inventory for Rio Cayapas, Northwestern Ecuador.
| Nombre común | Nombre científico | DMC (cm) | Gremio | Uso Comercial |
| Mazamorro | Aegiphila alba | 2 | 0 | |
| Alchornea | Alchornea | 3 | 3 | |
| Chirimoya | Annona? | 3 | 0 | |
| Guanábana | Annona? | 3 | 0 | |
| Guanábana arisca | Annona ? | 3 | 0 | |
| Guanama | Annona ? | 3 | 0 | |
| Peine mono | Apeiba aspera | 60 | 2 | 3 |
| alchorneoides | alchorneoides | 3 | 3 | |
| Chonta duro, chonta | Bactris gasipaes | 3 | 7 | |
| Banara | Banara ? | 2 | 3 | |
| Gualpité | Banara guianensis | 2 | 3 | |
|
Lirio,
lirio de monte |
Bombacopsis squamigera | 60 | 2 | 3 |
| Sande | Brosimum utile | 60 | 3 | 2 |
| Clavellín | Brownea multijuga | 60 | 3 | 1 |
| María | Calophyllum brasiliense | 3 | 1 | |
| Limoncillo | Calyptranthes sp. | 3 | 0 | |
| Sajo | Campnosperma panamense | 60 | 3 | 0 |
| Tangaré | Carapa guianensis | 50 | 3 | 5 |
| Cacadillo | Caryodaphnopsis theobromifolia | 60 | 3 | 5 |
| Piedrita | Casearia arborea | 2 | 1 | |
| Caucho | Castilla elastica | 40 | 2 | 6 |
| Cosedera, Guarumo, yarumo | Cecropia sp. | 60 | 1 | 0 |
| Cedro | Cedrela odorata | 50 | 2 | 4 |
| Ceibo | Ceiba pentandra | 60 | 2 | 2 |
| Pacora | Cespedesia spathulata | 60 | 3 | 0 |
| Caimitillo | Chrysophyllum argenteum | 60 | 3 | 0 |
| Nacedera | Citharexylum ? | 2 | 0 | |
| Nacedor | Citharexylum ? | 2 | 0 | |
| Nacedor | Citharexylum ? | 2 | 0 | |
| Moral bobo | Clarisia racemosa | 3 | 1 | |
| Moral | Clarisia sp. | 60 | 3 | 0 |
| Dormilón | Cojoba arborea | 50 | 3 | 1 |
| Compsoneura | Compsoneura sprucei | 3 | 0 | |
| Laurel | Cordia alliodora | 60 | 2 | 4 |
| Matapalo | Coussapoa sp. | 60 | 2 | 0 |
| Mortinio,capulí | Cybianthus sp. | 3 | 0 | |
| Pulgande | Dacryodes sp. | 50 | 3 | 2 |
| Dussia | Dussia | 3 | 0 | |
| Mambla | Erythrina poeppigiana | 40 | 2 | 6 |
| Guasca, guaso | Eschweilera sp. | |||
| Pepa sabrosa | Escweilera ? | 3 | 0 | |
| Sabroso | Escweilera ? | 3 | 0 | |
| Sabroso | Escweilera ? | 3 | 0 | |
| Eugenia | Eugenia ? | 3 | 0 | |
| Arrayán | Eugenia sp. | 3 | 0 | |
| Canalón, Costillo | Exarata chocoensis | 60 | 3 | 0 |
| Higuerón | Ficus máxima | 60 | 3 | 0 |
| Matapalo, higueroncillo | Ficus sp. | 40 | 3 | 0 |
| Madroño, madronio, madroño de monte | Garcinia sp. | 3 | 6 | |
| jagua | Genipa americana | 40 | 3 | 5 |
| Bolsa de toro | Grias peruviana | 3 | 6 | |
| Colorado | Guarea ? | 3 | 3 | |
| Pialde macho, pialde | Guarea kunthiana | 60 | 3 | 5 |
| Cargadera | Guatteria | 3 | 0 | |
| Guatteria sp. | 60 | 3 | 0 | |
| Paco | Gustavia sp. | 3 | 0 | |
| Cabo de hacha, huesito | Heisteria acuminata | 3 | 0 | |
| Carbonero, jubela, licania | Hirtella carbonaria | 3 | 0 | |
| Carrá | Huberodendron patinoi | 50 | 2 | 6 |
| Chanul | Humiriastrum procerum | 60 | 4 | 1 |
| Mascarey | Hyeronima alchorneoides | 50 | 2 | 1 |
| Guaba de mono | Inga ? | 3 | 0 | |
| Guabo1, guabillo | Inga latipes | 3 | 0 | |
| Guabo | Inga sp. | 3 | 0 | |
| Guabo tacuan | Inga sp. | 60 | 3 | 0 |
| Guabo machetón | Inga spectabilis | 60 | 3 | 6 |
| Pambil | Iriartea deltoidea | 3 | 7 | |
| Papaya monte | Jacaratia spinosa | 60 | 2 | 0 |
| Salero | Lecythis ampla | 60 | 3 | 1 |
| Wasdonguiichi | Leonia | 3 | 0 | |
| Poso, corazón embuelto | Maquira guianensis | 3 | 0 | |
| Matisia | Matisia | 3 | 0 | |
| Sapote | Matisia ? | 3 | 0 | |
| Sapotillo | Matisia malacocalyx | 3 | 0 | |
| Melastomataceae | Miconia | 2 | 0 | |
| Mora | Miconia sp. | 2 | 0 | |
| Mora, mora blanca | Miconia sp. | 2 | 0 | |
| Guayacán | Minquartia guianensis | 50 | 4 | 1 |
| Arrayancillo | Myrcia ? | 3 | 0 | |
| Hoja blanca | Naucleopsis ulei | 3 | 3 | |
| Canelón | Nectandra ? | 3 | 3 | |
| Jigua amarilla | Nectandra ? | 3 | 3 | |
| Jigua blanca | Nectandra ? | 3 | 3 | |
| Jigua negra | Nectandra ? | 3 | 3 | |
| Lauraceae | Nectandra ? | 3 | 3 | |
| Guadaripo | Nectandra guadaripo | 60 | 3 | 5 |
| Aguacatillo | Nectandra sp. | 60 | 3 | 3 |
| Jigua | Nectandra sp. | 40 | 3 | 5 |
| Balsa, boya | Ochroma pyramidale | 30 | 1 | 6 |
| Calade | Ocotea | 3 | 3 | |
| Aguacatón | Ocotea sp. | 60 | 3 | 3 |
| Calade | Ocotea sp. | |||
| Jigua pallante | Ocotea sp. | 60 | 3 | 3 |
| Chapil | Oenocarpus bataua | 3 | 7 | |
| Indio hediondo | Osteophloeum platyspermum | 60 | 3 | 2 |
| Cuángare, Sangre de gallina | Otoba gordoniifolia; O. gracilipes | 60 | 3 | 2 |
| Sapotolongo | Pachira aquatica | 3 | 0 | |
| Palicourea | Palicourea sp. | 3 | 0 | |
| Cuero de sapo | Parinari romeroi | 50 | 4 | 1 |
| Guión | Perebea guianensis | 3 | 0 | |
| Cabecita | Perebea xanthochyma | 3 | 0 | |
| Aguacate | Persea americana | 60 | 3 | 6 |
| Amarillo | Persea rigens | 60 | 3 | 3 |
|
Tagua |
Phytelephas aequatorialis | 3 | 7 | |
| Posoqueria | Posoqueria sp. | 3 | 0 | |
| Encanto, damagua, majahua | Poulsenia armata | 40 | 3 | 0 |
| Uvilla | Pourouma | 2 | 0 | |
| Uva de monte | Pourouma guianensis | 60 | 2 | 6 |
| Sapotaceae | Pouteria ? | 3 | 0 | |
| Caimito, caimitillo de monte | Pouteria sp. | 60 | 3 | 1 |
| Anime leña | Protium | 3 | 3 | |
| Anime |
Protium
sp. |
60 | 3 | 0 |
| Pisuhi, savaleta | Pseudobombax sp. | 60 | 2 | 3 |
| Frejolillo | Pterocarpus ? | 3 | 0 | |
| Bolsa de perro | Rauvolfia leptophylla | 3 | 3 | |
| Annonaceae | Rollinia ? | 3 | 3 | |
| Lechero | Sapium ? | 2 | 0 | |
| Sapium | Sapium ? | 2 | 0 | |
| Mata pez | Sapium sp. | 60 | 2 | 0 |
| Sloanea ? | Sloanea ? | 3 | 0 | |
| Sancona | Socratea exorrhiza | 3 | 6 | |
| Venenillo | Sorocea ? | 3 | 0 | |
| Quende, marequende | Sorocea sp. | 3 | 7 | |
|
Ovo |
Spondias mombim | 3 | 3 | |
| Huesito | Swartzia haugthii | 3 | 0 | |
| Machare | Symphonia globulifera | 50 | 3 | 1 |
| Tabebuia | Tabebuia | 3 | 1 | |
| Cucharillo | Talauma sp. | 50 | 3 | 5 |
| Guardián | Talisia sp. | 60 | 3 | 0 |
| Roble | Terminalia amazonia | 3 | 1 | |
| Barazón, Perdiz | Tetrathyllacium macrophyllum | 3 | 0 | |
| Cacao | Theobroma cacao | 3 | 6 | |
| Tovomita | Tovomita ? | 3 | 0 | |
| Tovomita | Tovomita ? | 3 | 0 | |
| Manglillo | Tovomita weddelliana | 3 | 0 | |
| Caraño | Trattinnickia aspera | 60 | 3 | 3 |
| Sapán | Trema integuerrima | 1 | 6 | |
| Pialdillo | Trichilia ? | 3 | 0 | |
| Cedrillo | Trichilia sp. | 60 | 3 | 3 |
| Cedro macho | Trichilia sp. | 60 | 3 | 3 |
| Chillialde, pichango | Trichospermum galeottii | 60 | 2 | 3 |
|
Turpinia |
Turpinia? | 3 | 0 | |
| Chalviandillo | Virola ? | 3 | 3 | |
| Myristicaceae | Virola ? | 3 | 3 | |
| Virola | Virola ? | 3 | 0 | |
| Chalviande, coco | Virola dixonii; V. reidii | 60 | 3 | 2 |
| Achotillo | Vismia ? | 2 | 0 | |
| Goma | Vochysia ferruginea | 2 | 3 | |
| Laguno | Vochysia macrophylla | 50 | 2 | 2 |
| Gualte | Wettinia quinaria | 3 | 7 | |
| Azafrán, tachuelo, cacho | Zanthoxylum riedelianum | 60 | 2 | 3 |
| Chípero, chispero | Zygia longifolia | 60 | 3 | 0 |
| Adulón | ||||
| Agua Agua | ||||
| Aguilla | ||||
| Ahuecador | ||||
| Aretero | ||||
| Bagatá | ||||
| Bagatá blanco | ||||
| Balsa macho | ||||
| Bambo | ||||
| Bantano | ||||
| Barbasco | ||||
| Barbasquillo | ||||
| Bejuco | ||||
| Bipupuchi | ||||
| Bombacaceae | ||||
| Cabezote | ||||
| Cacho de venado | ||||
| Cagua | ||||
| Candelilla | ||||
| Caracol | ||||
| Caracolillo | ||||
| Cascarillo | ||||
| Castaño | ||||
| Catanga | ||||
| Cauchillo | ||||
| Ceibo de papaya | ||||
| Chacarra | ||||
| Chanulillo | ||||
| Chiachi | ||||
| Chicharra | ||||
| Chijambi | ||||
| Chimbusa | ||||
| Chocho | ||||
| Chuncho | ||||
| Chundapuchi | ||||
| Clavo | ||||
| Culo de negra | ||||
| Cusumbi | ||||
| Desconocido | ||||
| Desconocido | ||||
| Dijchi | ||||
| Fabaceae | ||||
| Fruta de pajaro | ||||
| Guacharaco | ||||
| Guanamito | ||||
| Guatinero | ||||
| Guayabillo | ||||
| Guayabo | ||||
| Guayacanillo | ||||
| Hoja negra | ||||
| Jaboncillo | ||||
| Jaguilla | ||||
| Jeenchi**** | ||||
| Jeendachi**** | ||||
| Jentanjichi | ||||
| Jibon blanco | ||||
| Juvenchi | ||||
| Kijinuche | ||||
| Koyonipihuchi | ||||
| Kuguaichi | ||||
| Kuguayjinuche | ||||
| Kuhupugpuchi | ||||
| Kupalachi | ||||
| Kutupugpuchi | ||||
| Leguminosa | ||||
| Lengua de vaca | ||||
| Lengua de venado | ||||
| Llijcuipichi | ||||
| Macharillo | ||||
| Madronillo | ||||
| Manteca | ||||
| Mantecoso | ||||
| Marequende | ||||
| Masamorro | ||||
| Morocho | ||||
| Murunchi | ||||
|
Mutan |
||||
| Naranjo | ||||
| Narde | ||||
| Nipochi | ||||
| Pabo | ||||
| Pabo | ||||
| Pachaco | ||||
| Pacharaco | ||||
| Pacheco | ||||
| Pagnamuchi | ||||
| Palachi | ||||
| Paliarte | ||||
| Palmisha | ||||
| Palmito | ||||
| Palo blanco | ||||
| Palo pepa | ||||
| Palo santo | ||||
| Panecillo | ||||
| Pena de dios | ||||
| Pepa de mono | ||||
| Pepa de monte | ||||
| Piquelanchi | ||||
| Platano | ||||
| Popambuchi | ||||
| Quebra hacha | ||||
| Quelanchi | ||||
| Querre | ||||
| Quininande | ||||
| Rubiacea | ||||
| Sajillo | ||||
| Samia | ||||
| Sombra de rio | ||||
| Supachi | ||||
| Tambora | ||||
| Tanchachi | ||||
| Tegviachi | ||||
| Tejbiachi | ||||
| Tibijchiche | ||||
| Tsopa | ||||
| Tuchipiche | ||||
| Tyarinsa (palma) | ||||
| Vaina | ||||
| Vainilla | ||||
| Venandrillo | ||||
| Winula | ||||
| Zayvo |
Gremios:
1 Heliófitas efímeras
2 Heliófitas durables
3 Esciófitas parciales
4 Esciófitas totales
Uso comercial:
0 sin uso
1 madera dura para construcción( vigas, pisos, pilares)
2 madera de desenrollo y playwood
3 madera de encofrado y cajonería
4 maderas suaves (muebles, puertas, ventanas)
5 madera semidura
6 otros usos (frutos, látex, extractos, leña, fibras)
7 palmas