Abstract
This review provides an overview on the vegetation zonation of a large part of the mountain systems of the African continent. The Atlas and Jebel Marra are discussed as examples for the dry North African Mountains. The Drakensberg range is shown as representative of the mountain systems of Southern Africa. The main focus of the review falls on the Afrotropical mountains: Mt. Kenya, Aberdares, Kilimanjaro, Mt. Meru, Mt. Elgon, Mt. Cameroon, Mt. Ruwenzori, Virunga, Simien Mts., Bale Mts. and Imatong. A new nomenclature for the vegetation belts of Afrotropical Mountains is proposed.
Key words: Africa,
mountain vegetation, vegetation belts, nomenclature, Mt. Kenya,
Aberdares, Kilimanjaro, Mt. Meru, Mt. Elgon, Mt. Cameroon, Mt.
Ruwenzori, Virunga, Simien Mts., Bale Mts., Imatong, Atlas, Jebel
Marra, Drakensberg
Geology,
landforms and soil conditions
Africa
is an old continent. Nowhere else is so much of the surface covered
by basement rocks, and the larger part of the geology of Africa is
indeed the geology of pre-cambrian material (Petters 1991). Large
areas, so called cratons, have remained essentially unchanged since
the early Proterozoic (~ 2000 Mio BP). The mobile belts, i.e. the
huge basins and swells between these cratons, are composed of equally
old rocks, but have been subject to deformation and partly to
metamorphoses, mainly in the Pan-African orogenesis (700 - 500 Mio
years BP). Today, relief differences are relatively small in these
areas, so the continent can be described as a large, uneven plateau,
which is tilted to the northwest. Extensive erosion surfaces covered
by ancient, heavily leached soils are more widespread in Africa than
on any other continent, and the huge relatively flat Savannahs are
well known. Inselbergs are very characteristic remnants of the older
surfaces in these plains, but far too low to merit treatment here.
Equally well known as the flat surfaces are, however, some much more recent geological features, namely the huge, isolated volcanoes of the tropical part of the country, and the high fault mountains in the Atlas region and in South Africa. In these three regions, true high-altitude environments are found, with several ranges reaching well above 4000 m. These are almost exclusively located in the tropical parts, with Mt. Kilimanjaro (5985 m), Mt. Kenya (5198 m) and Ruwenzori (5109 m) being the highest sites. Several other mountains reach above the treeline, with the most extensive areas being in Ethiopia. However, the High Atlas in Morocco is also high enough to support truly alpine vegetation (4165 m).
Morocco occupies an intermediate position between the tectonic domains of southern Europe and Africa. The evolution is complex, and the atlas region was formed in several successive uplifting events. The south-westernmost part, the Anti-Atlas contains Precambrian material, but was largely formed in the late Proterozoic. Uplift occurred in Hercynian times in close correspondence to some mountains in Europe, and again mainly in late Cretaceous tectonic movements that formed the several chains of the High Atlas and the Saharan Atlas. The northern Riff and the Tell Atlas in northern Algeria are the southernmost outposts of the Alpine orogen, but recent Tertiary uplift affected also the older ranges, notably the Saharan Atlas. Most of the material is acidic, but several ranges display calcareous rocks, like the central High Atlas, part of the Saharan Atlas and the Rif. Only the High Atlas reaches above 3000 m, while the neighboring ranges hardly climb above 2000 m.
The central Saharan mountains Hoggar and Tibesti rest on the relatively stable West African craton. Part of the Tibesti is of ancient, Proterozoic origin, but there has been extensive recent volcanism. Both ranges are part of a chain of volcanoes of Cenozoic age that extends southwards to the central African Mt. Cameroon and eastwards to the East African Rift Valley System.
Mt. Cameroon is a huge single structure of largely Tertiary origin, but has been active ever since with more than 10 major events in the 20th century, some as recent as 1982 and 2000 (Ndam et al. 2001). The series of volcanoes starting at Mt. Cameroon has been described to continue up to the Jebel Marra in central Sudan. This is a massif rather than a single volcano, which has been active from the Miocene to the early Holocene (Miehe 1988). Lavas are predominantly basaltic in all these ranges.
Further eastwards, the huge volcanic plateau of Ethiopia rests on ancient material, but was largely formed in the tectonic processes during the formation of the East African Rift Valley system. The EARS is the prime example for a continental rift, and its huge structure extends over more than 6000 km from northern Mozambique along all of eastern Africa and through the Red Sea. Rifting started in the Cretaceous but has intensified in the Tertiary, and is most likely a consequence of tensional forces driving the two escarpments apart. Volcanism has been intensive and complex, following the two main branches, the Albertine Rift in the west, and the Gregory Rift in the east (Schlüter 1997).
The Ethiopian Rift is an extension of the latter, mainly formed by basaltic magmas in the Oligocene to Miocene, and reaching above 4000 m at several sites, notably in Simen Mts. in the northern part of Ethiopia and the Bale Mts. in the southwestern. Relatively deep and dry basins from the isolated volcanoes in east Africa proper separate this group of well-connected ranges.
Most of these are relatively young and of Pliocene to Quaternary age. This refers to mountains such as the Aberdares, which were formed from 3 Mio. BP onwards; Mt. Kenya was formed between 2-3 Mio. years BP (Heinrichs 2001); Mt. Kilimanjaro is even younger at 1-2 and Mt. Meru appears to be of similar age as are the Aberdare Mts. Lavas on the huge shields are predominantly alkaline or basaltic, but intermediate lavas formed several strato-vulcanoes.
The mineral composition is similar for Mt. Elgon, which is however an exception with respect to its age. Mt. Elgon lies at the border of Uganda and Kenya, and is thus not strictly located in the Rift Valley. Here volcanism commenced some 20 Mio years ago and lasted for about 10 Mio years. The neighboring Cherangani range is an ancient block of largely metamorphic rocks, but has been lifted to its present height in the Miocene. These two ranges have been high mountains, long before their neighbors in the eastern part of Africa, and also those in central Africa had attained high altitudes.
The Ruwenzori is a huge pre-Cambrian block, but has been lifted to alpine heights later than 2 Mio years BP. The Kahuzi-Biéga west of Lake Kivu has apparently experienced a similar evolution (Fischer 1996). The other high mountains along the Albertine Rift Valley are exclusively of volcanic origin, with volcanism commencing as early as some 13 Mio years BP in the Virungas. However, most of the activity occurred in the last 2 Mio. years and continued to the present, as was so strikingly demonstrated at the city of Goma, which was largely destroyed during a catastrophic lava outpour in 2001.
These young mountains are in striking contrast to the truly ancient Eastern Arc Mountains in Tanzania and northern Kenya. They are composed of granulite gneiss complexes and have been mountains during for over 25 Mio years. However, they barely reach 3000 m and lack an alpine belt typical for the other mountains discussed here.
Faulting during the formation of the Rift Valley has formed much of the present tropical part of African. Beyond Mozambique, mountains in the southernmost part of the continent are fault mountains like the northwestern Atlas system
The lower slopes of the volcanic ranges have been subject to intensive weathering and leaching, resulting in deep reddish soil profiles, mainly Acri- and Nitisols, occasionally Ferralsols. The dominant soil type of the upper montane belt are however well developed Andosols. The Andosols have moderately acidic soil pH values, and are exceedingly fertile and explain the intensive agriculture on the tropical mountains. The alpine regions are partly covered by Andosols, notably on the younger volcanoes, partly by various weakly developed soils on glacial deposits such as Regosols and Rankers. Very widespread are various Histosols that result from slow decomposition of organic matter due to coldness and stagnant water. They are characterized by low pH values (under 4 to 5.5) and generally low nutrient availability.
Altitudinal belts in the African
mountain regions
Vegetation belts in the North African
Mountains
Atlas
With
respect to their geologic origin, mountains in northwest Africa are
closely related to European mountains like the Alps. Correspondingly,
climate, flora and vegetation display strong similarities to
Mediterranean mountains like the Sierra Nevada in Spain, and mediate
to the arid sites of central northern Africa.There are three principal complexes, namely the High Atlas and its neighbors Anti-Atlas and Central Atlas; the Rif and Tell Atlas in northern Morocco and Algeria; and the Saharan Atlas in Algeria and Tunisia. These display large-scale climatic gradients, with conditions getting progressively drier with distance to the coast, so the Rif and Tell Atlas get most precipitation with annual totals above 750 mm in the northern foot zone. In the Moroccan Atlas, conditions get drier from west to east. These differences among ranges are small in comparison to the large differences between the northern and the south (-eastern) slopes within the mountain ranges. The northern lowlands receive well above 300 mm, while the southern slopes border the Saharan basin, where precipitation quickly drops to less than 100 mm in the lowlands (Weischet & Endlicher 2000).
Montane regions clearly receive more than lowlands with a precipitation maximum at 2000 - 3000 m elevation. This is around 750 mm in the High Atlas, and above 500 mm in all regions except for parts of the Saharan Atlas. The Rif and the Tell Atlas receive annual totals of above 1000 mm, which rise occasionally up to 2000 mm. Rainfalls occur almost exclusively in winter, so the summit regions are at least seasonally covered by snow, which might last until the following year in moist conditions.
Thus, moisture availability and the usual altitudinal temperature lapse rate control most of the vegetation patterns, but these are modified by edaphic components. Like in the Alps, calcareous and acidic bedrocks intersect, so vicariating plant communities are found depending on the geological background. This analogy is however limited, since the extremely harsh climatic conditions with a pronounced seasonality and the general dryness of the summit region induce excessive weathering. Thus the uppermost regions are covered with scree, which move in the frequent freeze-thaw cycles. Soils are weakly developed and conditions resemble those of dry ranges in the Sahara or even Central Asia, with an open and rather sparse vegetation cover completely different from the Alps.
Human impact has been tremendous in the entire region. Agriculture and livestock keeping were introduced some 5000 BP, and, like the entire Mediterranean region, most of the natural vegetation has been replaced by various secondary plant communities, rendering inferences of the potential character of the landscapes often different.
The Moroccan Atlas
Botanical exploration the area started equally late as in the seemingly much more remote tropical parts of Africa and started in the 1870's. It has however been intense and the general patterns are known since the middle of the last century (Rauh 1952). The High Atlas displays the typical situation. The northern foot zones are covered by open shrublands with Zizyphus ssp., Lotus and notably Acacia gummifera up to 900 m, where the dwarf palm Chamaerops humilis demarcates the transition to forest vegetation. Tetraclinis articulata forests are the natural community up to 1400 m, from where they are replaced by various communities dominated by Quercus ilex. Stands are dense and relatively shady with few companions as Cistus laurifolius on acidic, and Buxus sempervirens on calcareous bedrocks. Much of these forests have been replaced by various scrub communities with Pistacia lentiscus and Juniperus phoenicea in the lower regions, and by J. thurifera in the upper montane belt. Impressive Cedar (Cedrus atlantica) stands built a special forest community restricted to the moistest parts of the Atlas, but are more extensive in the Rif and in the Tell Atlas. Above 2800 m, J. thurifera forests dominate up to the treeline in some 3100 m. Unlike in other areas of Africa, progressively diminishing and finally shrubby growth of the dominant species do apparently not characterize the treeline ecotone. Instead, Junipers retain their single-stemmed growth up to the treeline, with stands opening progressively and distance between trees increasing.
Thorny cushion plants characterize the lower alpine region up to 3600 m. Cushions grow 0.5 to 1.0 m in height and have thick, often several meters long root. The dominant species are the widespread Alyssum spinosum, Bupleurum spinosum, joint by the flat thorny cushions of Arenaria pungens in upper elevations. In contrast, Erinacea pungens and Vellea mairei are restricted to calcareous bedrocks. Cushions prefer weakly inclined to level slopes where they can built almost closed stands, but have also benefited from anthropo-zoogenic human impact on the upper montane Juniper forest, where they from secondary stands on disturbed sites.
Moist sites in the lower alpine zone near small brooks are the habitat of communities called "Pozzines". These are meadows, physiognomically similar to alpine meadows in Europe, and thus colorfully flowering in summer. However, they are of very limited spatial extent, and are restricted to soils with impeded drainage over acidic bedrocks.
Substrate movement in the upper alpine region becomes increasingly strong above 3300 to 3600. The cushions become more and more restricted to the few level sites, and plants truly adapted to scree slopes and rock crevices take over. Again, we find vicariant taxa on acidic and calcareous substrates, like Viola dyris var. orientalis on calcareous rocks and Viola dyris var. typica on acidic bedrocks. The latter is an endemic taxon of the Atlas, other examples are Linaria lurida, L. heterophylla ssp. galioides and Vicia glauca var. rerayenssis, all growing on scree slopes. The relative importance of endemic species increases with altitude in the Atlas, and is around 30% in the lower alpine zone, while it reaches some 75% in the upper alpine and summit regions. There is no truly nival belt and plants occur up to the summit region.
The zonation on the southern slopes is somewhat different because of the greater aridity. The foot zone is covered by succulent Euphorbia species or open shrublands like on the northern slopes up to 1500. Juniperus phoenicea partly replaces Tetraclinis articulata and the belt of Quercus ilex forests is smaller and widely replaced by open Juniper scrub. Cedrus forests are entirely absent. The timberline is formed by J. thurifera as on the northern slopes, and the alpine vegetation is similar as well.
The Rif and the Tell Atlas
The Rif reaches barely 2500 m and the Tell Atlas is even lower, but affinities to the Moroccan Atlas are strong in the montane belt (Knapp 1973), so the description is kept short. The most widespread montane forest communities are also built by Quercus ilex, which forms dense and rather shady stands. Quercus suber forms extensive forests at the relatively moist coast of Algeria and Tunisia up to some 1000 m asl., but small stands of the Oak are also found in Morocco. Like Quercus coccifera, it has rather high moisture demands and is restricted to the coastal regions below 1200 m. Much of the areas have been cleared for agriculture, especially Olea europea plantations. The olive forms (semi-)natural stands with Pistacia lentiscus in drier parts of the lowlands, which might however be secondary in nature. Aleppo Pine (Pinus halepenis), often mixed with Quercus ilex, dominates much of the lower montane zone at 500 - 1500 m in the Rif and Tell Atlas, but is also occasionally found in the Moroccan Atlas. Cedrus atlantica forest is common in the moister ranges above 1500 m. However, the moistest sites there are covered by deciduous oak forests (e.g. Q. faginea) and coniferous forests with Abies maroccana and A. numidica. Juniperus phoenicea forests in the drier parts of the montane belt have suffered heavily from anthropo-zoogenic disturbance but occur up to 2400 m under natural conditions. Replacement communities include mixed scrub with various Cistaceae, Fabaceae and Ericaceae species in the lower montane belt, and shrublands with the dwarf palm Chamaerops humilis and Rosmarinus officinalis in the montane belt.
There is no truly alpine vegetation in the Rif and in the Tell Atlas simply because the ranges are too low.
The Saharan Atlas in northern
Algeria
The Saharan Atlas forms a second chain leeward of the Tell Atlas and is thus relatively dry. Much of the natural vegetation has been replaced by widespread steppes with Stipa tenacissima, that is an important species of secondary grasslands in the western Mediterranean region. It is accompanied by Lygeum spartum or Artemisia herbae-alba, mostly on dense or compacted soils. The natural montane vegetation closely resembles the situation in the Moroccan Atlas described above, notably in the western part of the country near the inter-state boundary. Communities with higher moisture demands are almost exclusively absent, and forest with Quercus ilex and occasionally Pinus halepensis would be the zonal vegetation of much of the northern slopes. The southern slopes are influenced by the arid Saharan climate and dry vegetation types with succulents climb up well into the montane belt. Where the mountains are high enough, alpine vegetation types occur, which are also similar to the High Atlas. Thorny cushions are relatively common, as are scree communities, but communities of moist rock crevices are impoverished and Pozzines are entirely absent. The alpine vegetation shows the typical differentiation of vicariating species depending on the presence of acidic or calcareous bedrocks as was described for the High Atlas.
Dry mountains in central Sudan - the
Jebel Marra
The
Jebel Marra is a highly isolated volcanic massif in the center of
Africa, more than 1500 km away from the nearest coast and more than
1000 km distant to the next mountain, with the Simen Mts. in northern
Ethiopia being the most "closely" related range. The Jebel
Marra emerges from a pedi-plain in 1000-1200 m, with a semi-arid
climate with down to 280 mm of annual precipitation (Miehe 1988). The
mountain slopes receive more rain, and the southwestern side is
clearly moister than the eastern. The belt of maximum precipitation
is situated at around 2000 m asl., where totals can be as high as
1800 mm in extreme years, but 800 mm appear to reflect the average
conditions better. For the summit region (max. 2976 m Times Atlas
3071) 600 mm have been estimated.
The vegetation shows correspondingly clear altitudinal gradients. Drought deciduous woodlands cover the piedmont and Acacia scrub, interspersed with lowland gallery forests along watercourses and in moist ravines and valleys, mainly in the southwest. These can be regarded as remote outposts of tropical evergreen forest. Typical species include evergreen Ficus ingens, F. thonningii, F. sur, Syzygium guineense and Trema orientalis, indicating affinities to tropical montane forest described below..
The dominant vegetation type in the submontane belt are, however, communities with Combretum molle and Boswellia papyrifera, which are replaced by wooded grasslands with Entada abyssinica and Acacia albida as a transitional belt to the montane communities.
The transition from these submontane lowlands to the montane, sparsely wooded grasslands occurs at some 2000 m. The principal tree species is Olea laperrinei, which forms riverine forests above 1900 m with Ficus palmata. The olive is also found in the Saharan mountains and indicates the relationships to Hoggar and Tibesti. However, the dominant vegetation are grasslands with Andropogon-Hyparrhenia, and Themeda triandra-Conyza hochstetteri. Scattered trees of O. laperrinei can be found on scree slopes and in the grasslands, indicating that at least part of these grasslands are suitable for tree growth. The upper montane grasslands above 2800 m are composed of Pentaschistis species and Festuca abyssinica. Occasionally, remnants of ericaceous vegetation, notably E. arborea, occur in the upper montane region. Ericaceous vegetation dominates the timberline ecotone in much of tropical Africa, but the Jebel Marra is not high enough for a true alpine belt.
Thus, with respect to the temperature and moisture availability most of the montane belt should be suitable for dry afromontane forests dominated by O. laperrinei, which have presumably been cleared by large-scale woodcutting, grazing and fire. This would clearly indicate parallels to the tropical mountains described in the following chapter.
Vegetation belts in the tropical
African mountain regions
Mount Kenya - A typical example for
the altitudinal zonation of tropical African mountain vegetation
Mt. Kenya
(Figure 1 a & b) is intermediate among the mountains in tropical Africa in
terms of geographical position and climate. Research there has
started early in the last century, so the available data are
relatively comprehensive. Hence, we will discuss the vegetation
zonation of Mt. Kenya in some detail, while descriptions for the
other mountains are kept shorter and focus on the differences to the
typical situation in central Kenya.Mount Kenya, located about 180 km north of Nairobi on the eastern side of the Great Rift Valley, is a widely isolated volcano. The base diameter is about 120 km; the northern foothills reach the equator. With its main peaks Batian (5198 m) and Nelion (5188 m), Mt. Kenya is, after Kilimanjaro (Kibo 5899 m), the second highest mountain in Africa. It is broadly cone-shaped with deeply incised, in the upper parts U-shaped valleys, indicating extensive former glaciations. Most parts of Mt. Kenya are still covered with forests up to approximately 3400 m in the South and 3000 m in the North, where a sharp boundary separates the forest from the lower alpine zone. Shape and location of the upper treeline has been severely influenced by fires. The present lower timberline is a result of extensive forest clearance and agriculture with cultivation reaching up to 1800 m on the southern, up to 2400 m on the eastern and western, and nearly up to 2900 m on the northern slopes. Without human impact, the mountain would be almost completely surrounded by dense forests.
As an important water catchment, Mt. Kenya contributes mainly to the Ewaso Nyiro and Tana River drainage systems, and is therefore of outmost importance for the water supply of about 50 % of the Kenyan population. Moreover, since other power stations did not fulfill the expectations hydropower from these rivers continues to generate more than half of the country's production of electrical energy (Berger 1985; Decurtins 1985, 1992; Leibundgut 1986). Densely populated farming areas surround the whole mountain, and large parts of the forest suffered heavily from encroachment in the last decades. The vertical zonation of the mountain has already been described in the early 20th century (Engler 1895; Allaud & Jeannel 1914), but the first comprehensive accounts are from the 1950s (Hedberg 1951).
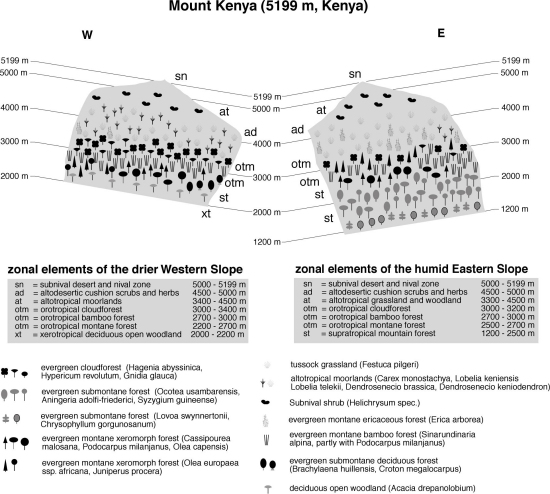
Fig. 1a. Vegetation zonation on Mt. Kenya, West-East Transect
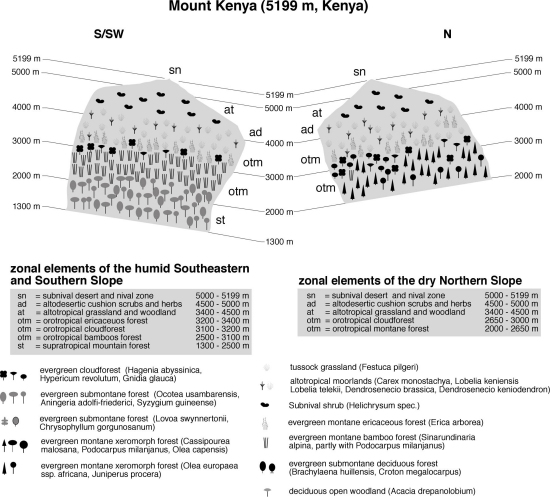
Fig. 1b. Vegetation zonation on Mt. Kenya, North-South Transect
Supratropical evergreen montane
forests
The
lowermost areas on the eastern and northeastern slopes from 1200-1500
m are covered with evergreen mountain forests, which closely resemble
forest types of central and western tropical Africa. Newtonia
buchananii, Lovoa swynnertonii and Chrysophyllum gorungosanum
are prominent tree species. These have survived only in
comparatively small remnants, while the principal forest formation
are typical Camphor forests, with Ocotea usambarensis, Aningeria
adolfi-friederici and Syzygium guineensis being the most
important canopy species. These evergreen submontane forests
(Bussmann 1994, 2001, Bussmann & Beck 1995a,b, 1999) are
two-storied, with huge specimens of the African Camphor tree, Ocotea
usambarensis in the upper canopy, and Xymalos monospora,
Lasianthus kilimandscharicus, Pauridiantha holstii and
Psychotria orophila in the lower tree layer and the
often-dense shrub stratum. Further characteristic species in the
upper tree layer are Strombosia scheffleri and Apodytes
dimidiata in the upper tree layer, while Tabernaemontana
stapfiana, Ochna insculpta, Macaranga kilimandscharica
and Peddiea fischeri form low trees and shrubs. Asplenium
sandersonii, A. elliottii and Panicum calvum occur
in the herb layer, together with Piper capense, Oplismenus
hirtellus, Plectranthus luteus, Begonia meyeri-johannis
and the ferns Dryopteris kilemensis and Blotiella
stipitata. Cyphostemma kilimandscharicum is a common
climber, and abundant fern epiphytes include Elaphoglossum lastii,
Trichomanes borbonica, Asplenium theciferum and
Oleandra distenta.On the lowermost eastern slopes of Mt. Kenya, forests with only one closed tree layer are found, dominated by Newtonia buchananii and many, often very tall palms (Phoenix reclinata), which give these areas an appearance like an Andean cloud forest. Once, these forests covered large tracts of land especially on the eastern slopes of Mt. Kenya, and formed the lowermost submontane forest belt in transition to the savannah lowlands, but these forests have long ago been cleared for cultivation.
On the western side of Mt. Kenya, evergreen submontane semi-deciduous forests, where drought resistant species like Calodendron capense or Croton megalocarpus are common, follow woodlands dominated by Acacia drepanolobium from 2000-2300 m.
On the northern part of Mt. Kenya, the woodlands lead directly into very drought-resistant xeromorphic forests, which are almost entirely dominated by Pencil Cedar (Juniperus procera) and Wild Olive (Olea europaea subsp. africana). A Bamboo belt is absent there, and even the narrow cloud forest belt is heavily interspersed with Cedar. Several authors mentioned a so-called "forest gap" on the drier northern slopes of the mountain (e.g. Hutchins 1909; White 1950). Whether this gap is natural or man made has been a matter of debate. According to the statements of long-term residents of the area, clearing the forest for farming purposes has widened the gap. In the late 1970's, the Kenyan government for farmland excised some areas of the Mt. Kenya Forest Reserve. In many smallholder fields (shambas) of this area, forest trees or remnants of trees are still present, indicating clearly the former extension of the forest belt. Climatic conditions are less probable to be responsible for the gap since in an even drier area exactly north of the gap on Mount Kenya, the Ngare Ndare forest with extensive stands of Juniperus procera is found. Therefore, there are no reasons to assume a natural reason for the existence of the forest gap. Most probably, a formerly closed forest belt must be concluded.
Riverine forests and a variety of very different forests can be also found, indicating affinities to the mountain forest communities of Central and Western Africa. Lovoa swynnertonii, a very tall and dominant canopy tree is a typical species, growing together with the shrubs Rawsonia lucida, and with Heinsenia diervilleoides and Rinorea convallarioides. In many places Uvariodendron anisatum, with its beautiful and very fragrant white leaves, grows abundantly in the lower tree stratum and in the shrub layer.
Pure Camphor forests are found mainly on the very wet southeastern and southern slopes of Mt. Kenya on altitudes between 1550-2550 m. They grow on humic Niti- and Acrisols and receive an annual rainfall of 1500-2500 mm. Moist forests in the lower and middle submontane region of southeastern Mt. Kenya are dominated by the evergreen species Syzygium guineense (Myrtaceae) and Aningeria adolfi-friederici (Sapotaceae), reaching a height of up to 50 m. Furthermore, the shrubs Drypetes gerrardii (Euphorbiaceae) and Allophyllus cuneatus (Sapindaceae), as well as the lianas Adenia gummifera (Passifloraceae) and Jaundea pinnata (Connaraceae) are characteristic species. The number of epiphytic mosses is limited, but a wide variety of epiphyllous and epiphytic liverworts and lichens occur.
Most stands suffered from heavy exploitation, and have been replaced by secondary vegetation types, leaving untouched evergreen forests only in steep ravines or in remote areas, which are difficult to access. Macaranga kilimandscharica dominates the canopy of these secondary forests, and often forms dense pure stands. It is a very fast growing species suppressing the regeneration of other trees.
Orotropical montane forest - Evergreen
broad-leaved and evergreen xeromorphic montane forests
The
cedar forests (Juniperus procera) grow on humic Acrisols
(Speck 1986) at rainfall totals between 700 and 1500 mm. They
represent the typical vegetation of the altitudinal range between
2500 and 2950 m of the drier exposures (Bussmann 1994, 2001, Bussmann
& Beck 1995a, 1999). Evergreen-broadleaved forests cover the
moister southern and southeastern slopes, at lower altitudes between
2150-2650 m. Juniperus procera itself is rare or completely
absent, due to heavy logging and suppressed regeneration in the
closed stands. The dominant tree species in these two-storied stands
are Cassipourea malosana (Rhizophoraceae) and Olea capensis
subsp. hochstetteri, the East African Olive; both forming the
upper canopy. Lepidotrichilia volkensii in the lower tree and
shrub strata, and Ilex mitis, the African Holly are further
important woody species.Cedar forest, in contrast, is rather open and one or two-storied, with J. procera growing about 50 m tall under favorable conditions. Other important tree species, mainly of the lower canopy, are the Wild Olive (Olea europaea subsp. africana) and Podocarpus latifolius. Mosaics of dense grass layers with Stipa dregeana and Brachypodium flexum, interspersed with herbs (Sanicula elata, Isoglossa gregorii and Achyranthes aspera), and low shrubs (e. g. Berberis holstii) are also characteristic of these open cedar forests.
Orotropical Bamboo forest
Dense
forests dominated by the African Bamboo (Sinarundinaria alpina)
which are found on various East African mountains, constitute a very
distinct formation (Bussmann 1994, 1997, Bussmann & Beck 1995a,
1999, Bussmann 2001, Bytebier & Bussmann, 2000). Sinarundinaria
alpina is the overwhelmingly dominant species in all strata, but
Impatiens hoehnelii, Pseudocarum eminii as well as
Selaginella kraussiana, Cyperus dereilema, and
Anthriscus sylvestris are common companions. Bamboo forests
are restricted to a relatively narrow ecological range, mainly
depending on soil temperatures of 10-15 °C and the presence of
very deep volcanic soils, namely humic Andosols. On Mt. Kenya, they
are found in an almost closed belt around the entire mountain,
interrupted only on the dry northern slopes. Stands are somewhat more
extensive on the very wet southern parts.On steep slopes of the western, northwestern and eastern side of Mt. Kenya, in areas with lower rainfall and higher soil temperatures, often huge individual trees protrude from the dense Bamboo layer in some 10-15 m height. The dominant species of the tree stratum is always Podocarpus latifolius, which is also common in the surrounding montane forest. Towards the hygric limits of the Bamboo, mainly on the dry northwestern parts of the mountain, Juniperus procera and Olea europaea subsp. africana are found as characteristic members of the tree stratum. Trails and heavily trampled resting places of big game occur everywhere in these forests, and seem to facilitate regeneration of trees in the otherwise extremely shady stands of Sinarundinaria alpina.
Orotropical cloud forest
Two forest types, namely tall and malacophyllous forest with Hagenia abyssinica occupy this vegetation zone, and lower stands built by various species with sclerophyllous, "ericoid" leaves. The so called "ericaceous belt" forms the transitions zone towards truly afroalpine vegetation within the timberline ecotone. In contrast, the malacophyllous forests extend from 2900 to 3300 m a.s.l., where low clouds and mist are frequent. The dominant tree is the Kosso tree, Hagenia abyssinica (Rosaceae), often accompanied by the Giant St. John's Worth, Hypericum revolutum. Bothriocline fusca is a common shrub, while Polygonum afromontanum, Stephania abyssinica, Cinereria deltoidea and Carduus afromontanus are common herbs. Pure Hagenia-Hypericum forests with a low, dense shrub layer of Hypericum are characteristic for the wetter southeastern to western slopes of Mt. Kenya. Here, important herbaceous companions are Luzula johnstonii, the large Lobelia bambuseti, Rubus frieseorum, Agrostis schimperiana, Poa schimperiana, Helictotrichon milanjeanum, and one of the few succulents, Uebelinia rotundifolia.
In the drier northern part of Mt. Kenya, the Hagenia-dominated forests are replaced by mixed forests with Podocarpus gracilior and broad-leaved species, clearly differentiated by the high frequency of Juniperus procera, Olea europaea, Nuxia congesta and especially Rapanea melanophloeos. Hagenia abyssinica is only a co-dominant canopy species and Hypericum revolutum is mostly absent.
From some 3300 m upwards, open ericaceous communities gradually replace Hagenia forests. They grow at the upper limit of forests, but the timberline has been lowered by several hundred meters due to fires and undulates now at 3100 to 3300 m. True Erica excelsa forests are poorly developed on Mt. Kenya, and most often the ericaceous belt is formed by remnant stands of ericaceous scrub with Erica trimera (formerly Philippia, Oliver 1987), E. arborea, Conyza vernonioides, and the astonishingly "ericoid" Asteraceae Stoebe kilimandscharica (Rehder et al. 1988). The latter, together with Protea kilimanscharica often indicates regular disturbance by high altitude fires. Fires are usually discontinuous, so the scrub is interspersed with various grassland types that are already afroalpine. Thus, ericaceous and afroalpine communities generally form patchy mosaics and transition communities rather than clear altitudinal belts.
Altotropical grasslands and
moorlands
In
tropical Africa, the altitudinal belt above the timberline has been
termed "afroalpine" (Hauman 1955). Communities intersect
with ericaceous vegetation, and are mainly dominated by tussock
grasses and stands of Giant Rosette Plants. Fortunately, a vegetation
map is available, so the spatial distribution of communities is well
known (Rehder et al. 1988; Beck et al. 1990). "Open Moorlands"
are particularly extensive on the western slopes of Mt. Kenya at
altitudes from 3400-3800 m, often found at clearings of ericaceous
vegetation. The dominant species are grasses, namely the large sedge
Carex monostachya, interspersed with tussocks of Festuca
pilgeri in drier places. Most characteristic are the large,
cabbage-like ground rosettes of Dendrosenecio brassica and
Lobelia keniensis. The Giant Rosette "tree"
Dendrosenecio johnstonii subsp. battiscombei is common
in the lower moorlands as well as in remnant ericaceous stands.
The "Upper Moorland Zone", extending from about 3800 m upwards as high as 4550 m comprises two very characteristic vegetation units. Where moisture availability is sufficient, "Upper Alpine Wetlands" are formed by Dendrosenecio brassica and Lobelia deckenii ssp. keniensis. In contrast to lower altitudes, sclerophyllous dwarf shrubs like Alchemilla johnstonii, Alchemilla argyrophylla subsp. argyrophylla and Helichrysum brownei often replace grasses. Sedges are less important than in the lower moorlands.
The typical plant community above 4300 m is the "Dendrosenecio woodland", in which D. brassica is replaced by the Giant Groundsel, Dendrosenecio keniodendron, growing together with Lobelia telekii. From 4500 m onwards, the Giant Groundsels recede, and an open layer of Festuca pilgeri, interspersed with Lobelia telekii, Festuca abyssinica, Carduus chamaecephalus and Senecio keniophytum extends to the subnival zone. Occasional specimens of vascular plants where found up to the summit region in above 5000 m (Rehder et al. 1988). In contrast to the situation on Kilimanjaro, where low precipitation creates a real alpine desert, closed vegetation on Mt. Kenya is more limited due to freezing and continuous solifluction, although moisture availability probably has some influence (Beck 1994).
Mt. Kenya's neighbors: Mt. Kilimanjaro,
Mt. Meru, Aberdare Mts. and Mt. Elgon
Although
they are separated by hundreds of kilometers of lowlands, the
neighboring mountains show a vegetation zonation very similar to Mt.
Kenya. These striking similarities have long been known to
phytogeographers (Hedberg 1951; Hedberg 1964), and have let to the
general classification scheme that provided the base for the previous
chapter. The vegetation belts are very distinct, as are differences
between sides of the mountains, where drought-resistant forest types
on their northern escarpments replace very humid elements on their
southern and southwestern slopes.
Exposure differences are particularly pronounced on Mt. Kilimanjaro (Figure 2). The southern slopes are unusually moist with an annual precipitation of around 900 mm in the southern foothills, around 2000 mm at 1500 m asl. and well above 3000 mm between 2000 and 2300 m. The summit region is, in sharp contrast, an alpine desert with well below 200 mm annual precipitation (Hemp 2002). Although no exact measurements are available, the northern slopes are clearly drier as indicated on the available vegetation map and the accompanying descriptions (Hemp 2001). Below 1000 m dry savannah surrounds the entire mountain, but on the northern slopes savannahs with Acacia drepanolobium and forest plantations climb above 2000 m altitude, followed by a small belt of xeromorphic forests with Cedar and Olive up to 2800 m.
In the moist south between 1000 - 1800 m, evergreen submontane deciduous forests has largely been replaced by dense agriculture. Remnants of the lowermost submontane forests are characterized by Olea europaea and at drier sites by O. capensis, but from 1700 m upwards, lower montane evergreen Camphor forests and other broadleaved forests dominated by Cassipourea malosana replace these. In proper montane forests above 2000 m Podocarpus latifolius becomes increasingly common. The most striking feature of these forests is the abundance of epiphytes, in particular Hymenophyllaceae and other fern groups, which give the stands the appearance of a true elfin forest. On all sides of Kilimanjaro, dense Erica excelsa forests occupies the belt between 2700-3100 m (South) and 2800-3400m (North). Lower down, in the central montane belt, such forests form replacement communities at disturbed sites (Hemp & Beck 2001). Cloud forests with Hagenia abyssinica and Hypericum revolutum are restricted to special sites such as boulder streams and moist valleys. Above 3100 m, Erica forests are gradually replaced by ericaceous scrub mixed with altotropical moorlands, where subspecies of Dendrosenecio johnstonii and Lobelia deckenii grow as vicariant taxa to Mt. Kenya (Knox 1993). Above 3900 m, subnival Helichrysum scrub takes over up to some 4600 m, where it becomes to dry for growth of vascular plants. The summit region is truly nival and devote of vegetation.
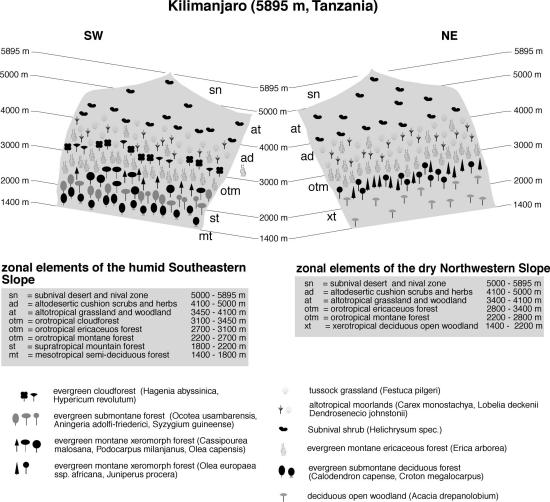
Fig. 2. Vegetation zonation on Mt. Kilimanjaro
The northern flanks of the neighboring Mt. Meru (Figure 3) are covered by deciduous open woodlands, which are followed by a small band of cloud forest with Hagenia abyssinica, Hypericum revolutum and Gnidia glauca between 2500-3000 m (Hedberg 1955; Hedberg 1964?). Higher up, these are replaced by evergreen ericaceous scrub up to 3700 m, which forms the transition to altotropical moorlands, where Carex monostachya, Lobelia deckenii and Dendrosenecio johnstonii dominate. On the southern slopes of this mountain, however, evergreen montane xeromorphic forest with Podocarpus latifolius, Cassipourea malosana and Olea capensis grows at altitudes from 1700-2300 m. These merge gradually into a distinct Bamboo belt that is replaced by typical cloud forest at 2550-2700 m. The zonation of the uppermost regions shows the same features as the northern escarpment.
The Aberdare Mts. form the next neighboring mountain range west of Mt. Kenya. Climate and vegetation were comprehensively described by Schmitt 1991, who also provides a vegetation map. Climatic conditions are similar to Mt. Kenya, with highest precipitation in the southeast and a maximum in the montane belt of around 2200 mm. The vegetation zonation is also largely similar, with Cassipurea malosana and Olea capensis in the supratropical mountain forest. Above 2500 to 2700 m, forests with Cedar, Podocarpus latifolius and riverine forests with Afrocrania volkensii dominate. An almost closed belt of Bamboo, from which specimens of P. latifolius and Nuxia congesta emerge, follows these. Above 2900 m cloud forests with Hagenia abyssinica, Hypericum revolutum and Rapanea melanophloeos take over. Erica excelsa becomes important towards the upper limit of montane forests on the main plateau in some 3100 m altitude. Thus, most of the upper region of the Aberdare Mts. is well in the altitudinal range of ericaceous forests and scrub, so that grasslands are largely secondary and promoted by fires as well as grazing by wild ungulates. The vegetation gives a very patchy impression, and altitudinal belts can hardly be inferred except towards the summits from 3600 m upwards. Ericaceous vegetation includes well-developed cloud forest of E. excelsa, and scrub with E. trimera, Stoebe kilimanscharica and Cliffortia nitidula. The altotropical grasslands are dominated by Festuca pilgeri, Koeleria capensis, the C4 grass Andropogon lima, and at moist sites sedges (Carex monostachya, Cyperus kerstenii). Again, vicariant subspecies of the Dendrosenecio johnstonii and of the Lobelia deckenii groups grow in the afroalpine zone.
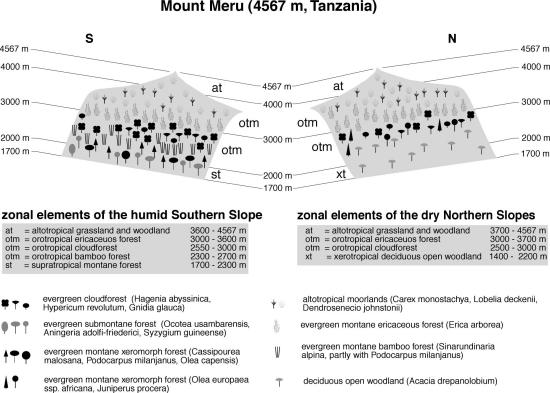
Fig. 3. Vegetation zonation on Mt. Meru
Near the boundary to Uganda, Mt. Elgon and the Cherangani Mts. form the easternmost part of these semi-humid mountain ranges. The Cheranganis are heavily cultivated and grazed on all elevations, so there is hardly any natural montane forest left. With some 3600 m altitude they are not high enough for extensive natural grasslands, so much of the present altotropical grasslands appear to be secondary (Mabberley 1975).
In contrast, Mt. Elgon is the largest solitary volcano with a full set of altitudinal belts comparable to Mt. Kenya (Wesche 2002, vegetation maps provided by van Heist 1994a; van Heist 1994b; Neville & Wesche in press). The western and southwestern slopes are clearly moister than the eastern part of the mountain, with a maximum of precipitation between 2300 and 2700 m asl. (1500-2000 mm on the western slopes, 1200-1500 on the eastern). Hardly any natural vegetation has survived the intense agriculture below 2200 - 2500 m, but higher up, large supratropical forests with scattered Camphor, Aningeria adolfi-friederici, Podocarpus latifolius and riverine Afrocrania forests survived. The eastern side is covered by dry orotropical forest with P. gracilior, Olea capensis and scattered Cedars. The Bamboo belt is almost closed except in the northernmost part. Cloud forests with Kosso and tree heather have mainly survived in the western part. Above the present timberline, mosaics of ericaceous vegetation with E. trimera and Stoebe kilimandscharica, Helichrysum and Alchemilla dwarf scrub and altotropical Festuca pilgeri grassland form the vegetation up to some 3700 m. Above this, afroalpine grasslands, Helichrysum scrub and Dendrosencio woodlands extend up to the summits. Mt. Elgon carries a separate species within the Dendrosenecio johnstonii complex, with one subspecies occurring up to 4000 and a second one between 4000 and 4300 m, resembling D. keniodendron on Mt. Kenya.
The interlacustrine highlands:
Ruwenzori, Virunga Volcanoes and Kahuzi-Biéga
The
mountains at the western branch of the Rift Valley form a separate
group due to their perhumid rather than semi-humid climate. They
border the huge rainforests of the Congo basin, so their lower slopes
are covered by luxurious tropical evergreen forest with a high
richness in species. Comprehensive vegetation surveys have been
produced for Kahuzi-Biéga (Fischer 1996, including land cover
map), the eastern half of the Ruwenzori range (Schmitt 1992; Osmaston
1996, land cover map by van Heist 1999, glaciological map by Osmaston
& Kaser 2001), and for parts of the Virunga Mts. ((Snowden 1933;
Biedinger 1995; Karlowski 1995).
Although the eastern side is generally drier, the vegetation of Ruwenzori (Figure 4), as of Mt. Cameroon, shows no clear distinction between the humid Western and drier Eastern side of the mountain, whereas on the Virunga Volcanoes (Figures 5 and 6) exposition differences are pronounced. The evergreen submontane forest belt of Ruwenzori stretches from 1000-1600 m. Common and typical tree species are Ocotea usambarensis, Aningeria adolfi-friederici and Syzygium guineense. On Muhavura and Karisimbi, submontane forest occupies both western and eastern slopes from 1300 to 2000 m. At altitudes between 2000-2300 m evergreen supratropical mountain forests with Olea capensis, Cassipourea malosana and Podocarpus latifolius replace lowland species in all three regions. In striking contrast to East Africa, no Cedar (Juniperus procera) can be found on any mountain in central Africa. Bamboo (Sinarundinaria alpina), mostly in stands interspersed with P. latifolius, is a prominent feature on Ruwenzori (from 2300 - 3000 m) and on the Virungas (2400-2600 m on Karisimbi, and stretching up as high as 2900 m on western Muhavura).
On Ruwenzori, evergreen Erica forest follows immediately after the Bamboo belt and forms extensive stands at 3000-3900 m. The situation on the Virunga Volcanoes is different. On Western Karisimbi (Figure 5), a small fringe of evergreen cloud forest with Hagenia abyssinica and Hypericum revolutum separates Bamboo and ericaceous belt between 2600-2700 m, from which ericaceous forests can be found up to 3800 m. The cloud forest belt is better developed on the humid eastern slope, where it extends from 2600-3500 m, followed by ericaceous scrub up to 3800 m. The drier Muhavura (Figure 6) shows an exactly inverted cloud forest distribution: on the more humid western slope, Hagenia prevails from 2900-3300 m, followed by the ericaceous vegetation up to 3700 m, whereas on the dry eastern escarpment the cloud forest zone is completely replaced by afroalpine grassland (Karlowski 1995).
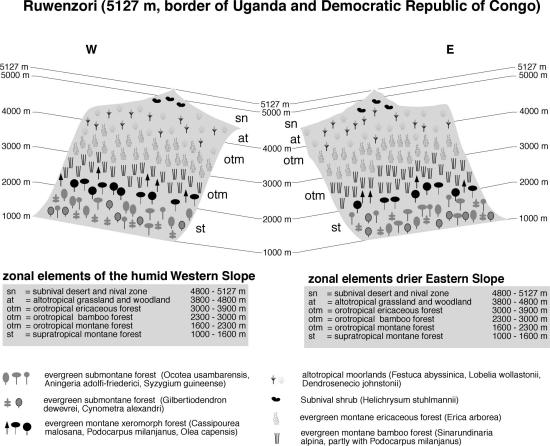
Fig. 4. Vegetation zonation on Mt. Ruwenzori
Extensive altotropical moorlands always occupy the alpine zone of the Central African Mountains. Tussocks of Festuca abyssinica and Carex runssoroensis, as well as the giant rosette plants Lobelia wollastonii and Dendrosenecio johnstonii occur on all three mountains, whereas Lobelia stuhlmannii grows only on the Virungas. Due to its much higher elevation, upper alpine dwarf scrub is only found on the uppermost parts of Ruwenzori, where Helichrysum stuhlmanii forms isolated thickets. The Kahuzi-Biéga range is not high enough for a true afroalpine zone, so the summit region is covered by ericaceous scrub and Deschampsia flexuosa grassland. The latter indicates the regular presence of fires.
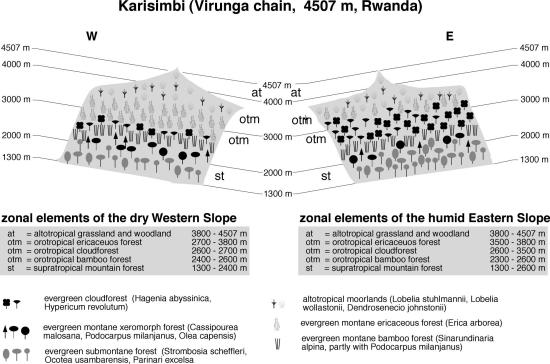
Fig. 5. Vegetation zonation on Mt. Karisimbi, Virunga Volcanoes
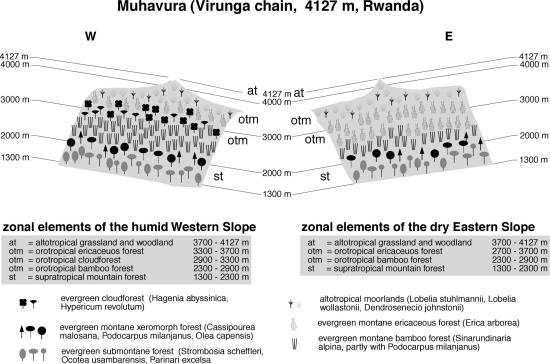
Fig. 6. Vegetation zonation on Mt. Muhavura, Virunga Volcanoes
The westernmost outpost - Mt.
Cameroon
The
highlands in Cameroon form a widely isolated archipelago in western
central Africa. The highest peak is Mt. Cameroon (4095 m, Figure 7),
an active volcano that had its most recent eruption in 1982. The
climate is extraordinary moist, with up to 10000 mm annual
precipitation in the foot zone and below 2000 mm in the summit region
(Cable & Cheek 1998, including vegetation map). Mean air
temperature is around 20°C at 900 m and estimated to be around 4°
in the summit region. Snow is rare, even on top of Mt. Kilimanjaro.
Soils in the mountains are mostly young and therefore fertile, so
large areas have been cleared for agriculture. About a third of Mt.
Cameroon is now under legal protection, and here natural vegetation
has survived partly down to the sea level (Cable & Cheek 1998).
The small remnants of lowland forests are rich in various species,
and have the highest number of endemics of all vegetation belts (29
vs. 49 for the entire range). Pluviotropical evergreen rain forest
dominate the larger part of these altitudes, with tree species like
Schefflera mannii or Syzygium guineense frequently
found in the canopy, and Allophyllus bullatus forming a second
layer. These forests merge gradually into submontane evergreen forest
from 800 m upwards. Species composition, however, changes only
slightly, being characterized by Syzygium staudtii,
Schefflera abyssinica, and still Allophyllus bullatus
(Richards 1963; Hall 1973). From 2000-2600 m orotropical cloud
forest is the dominant forest type with species found on all tropical
African mountains like Nuxia congesta, Prunus africana,
Xymalos monospora, and Hypericum revolutum. These form the
evergreen cloud forest, together with Rapanea neurophylla.
Bamboo is completely absent from Mt. Cameroon, but occurs other
ranges in western central Africa. Hagenia abyssinica does also
not occur on Mt. Cameroon, although all other species show clear
relationships to the Hagenia-dominated upper montane forests
common in other areas of sub-Saharan Africa. At all altitudes, large
parts of the forests have been replaced by grasslands due to high
volcanic activity and thus frequent fires. For the same reason, scrub
and thickets are frequently found all over the mountain, with
Marantaceae dominating below 2000 m and ericaceous species like Erica
mannii, Agauria salicifolia and Myrica arborea forming
a patchy ericaceous belt towards the timberline ecotone. Dense
altotropical tussock grasslands reach up to 3400 m, where large
Poaceae as Andropogon sp., Festuca abyssinica, Deschampsia
mildbreadii and Loudetia camerunensis dominate,
interspersed with small tree islands (Photo). True giant rosette
plants, Dendrosenecio or Lobelia species are absent
from Mt. Cameroon, though Crassocephalum mannii and Lobelia
columnaris show resemblances to this growth form. From 3400 m
upwards to the summit region, vegetation cover becomes more open and
impoverished in species (Maitland 1932). There are no clear
vegetation differences with regard to slope exposition.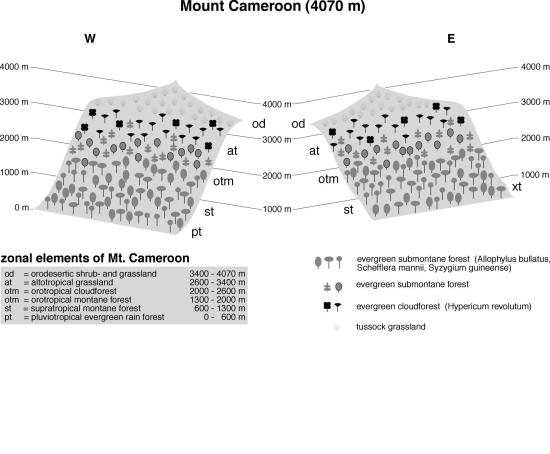
Fig. 7. Vegetation zonation on Mt. Cameroon
The Sudano-Ethiopian Mountains:
Imatong, Simien, Bale
In
striking contrast to the mountains in the strictly tropical part
Africa, the Sudano-Ethiopian mountains emerge from dry vegetation and
semi-arid surroundings. Thus, the mountains of southern Sudan and
Ethiopia are characterized by steep climatic gradients and
correspondingly, by clear altitudinal changes in the vegetation
zonation. All ranges are widely isolated from the eastern African
mountains, with the Imatong Mts. in southern Sudan being the least
distant.Their foot zone receives some 800 mm, while at 1900 m asl. some 2200 mm have been recorded; the highest figure in Sudan. The lower slopes of the Imatong Mts. (Figure 8) are covered with lush evergreen submontane forest, where Ocotea usambarensis, Olea welwitschii and Chrysophyllum fulvum are common species (Jackson 1956). Above 2000 m, these forests merge into evergreen montane xeromorphic forest with Podocarpus latifolius and Olea capensis. Above 2600 m, a mixture of evergreen Bamboo and evergreen cloud forests with Hagenia abyssinica replaces these. The highest zones, above 3000 m, are covered with dense thickets formed by Erica arborea and Myrica salicifolia up to the summit region.
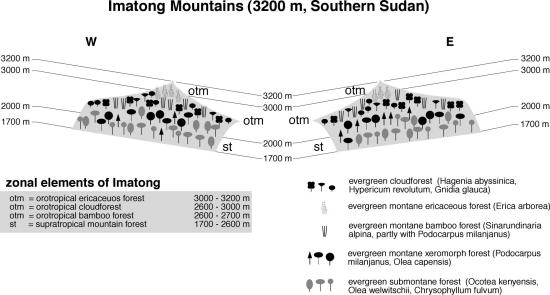
Fig. 8. Vegetation zonation in the Imatong Mts.
The by far largest area of afromontane and afroalpine environments is found in Ethiopia. This huge country displays notable differences in climate, and consequently relatively moist ranges as the Bale Mts. in the southern part are found, as well as extremely dry areas like the Simen Mts. in the north. The Bale Mountains (Figure 9) form the single largest afroalpine area, with a flora and vegetation being transient between East Africa and the drier northern part of Ethiopia (Weinert 1983). The southern foot zone emerges from dry Combretum-Commiphora savannah at 390 mm annual precipitation, but the southern Harenna escarpment receives around 850 mm at 3000 m asl. The central part of the range is a huge plateau in some 4000 m asl., where conditions become progressively moister towards the northern slope, which receives around 1000 mm (Miehe & Miehe 1994). This induces a clear zonation of very well pronounced vegetation belts. The lower southern slopes are covered with dense evergreen submontane deciduous forest from 1450-1900 m, where Podocarpus gracilior, Filicium decipiens and Celtis africana as well as C. gomphophylla form an open canopy (Lisanework & Mesfin 1989; Bussmann 1997). The undergrowth is often dominated by very dense thickets of wild coffee (Coffea arabica). Mid-altitudes from 1900-2300 m are covered by evergreen submontane forest (with Ocotea kenyensis, Aningeria adolfi-friederici and Olea welwitschii as most characteristic tree species), and thus resemble the Imatong Mountains closely. Above 2300 m, Cassipourea malosana, Schefflera abyssinca and Croton macrostachyus form the evergreen montane forest, which slowly gives way to cloud forest communities with Hagenia abyssinica and Hypericum revolutum from 2400 m onwards. Large areas of Bamboo are found between 2800-3100 m altitude. Cultivation has been more intense on the drier northern slopes, so only remnants of the natural vegetation survived. Dry orotropical Cedar and Podocarpus forests have been described up to 3000 m (Weinert 1983), but cloud forest with Kosso and Giant St. John's Worth is restricted to a band from 3000-3400 m.
Ericaceous vegetation with E. trimera sl. start to dominate the vegetation from this zone upwards to 4000 m. As elsewhere, the vegetation of the ericaceous belt has been severely altered by human influence, so mosaics of forests, scrub and afroalpine communities result. On the central Sanetti plateau, woody vegetation is restricted to sheltered sites as outcrops or boulder streams up to 4 up to 4000m. Instead, Helichrysum scrub and to a lesser extend tussock grasslands prevail up to the highest peaks (Menassie & Masresha 1996). Many of the grass and shrub species demonstrate clear affinities to the eastern African Mts. (e.g. Pentaschistis pictigluma, Helichrysum citrispinum), but the Giant Lobelias are present with only one endemic species (Lobelia rhynchopetalum), and the widespread forest species Lobelia gibberoa. Giant Groundsels are not found in Ethiopia. Strangely, an endemic thistle (Echinops longisetus) has developed a stem here resembling giant groundsels.
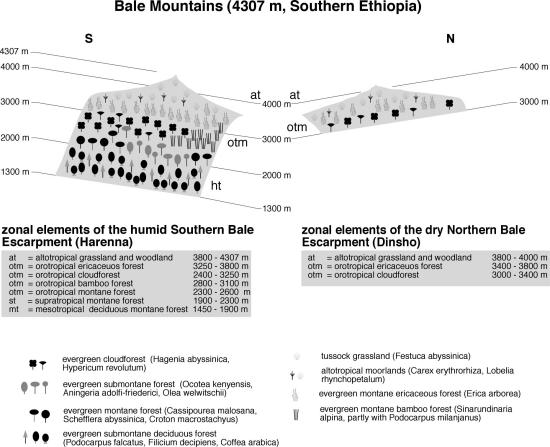
Fig. 9. Vegetation zonation of the Bale Mts.
Much of the Ethiopian highlands would bear montane forests if untouched, hence remnants of these forests still occur in the central part of the country (Tamrat 1993). The plant communities are still strikingly similar to those described for Mt. Kenya, but differences increase north-westwards, until the dry Simen Mts. (Figure 10) form the second largest afroalpine region in Ethiopia. Here, the precipitation regime finally becomes unimodal; the foot zone is rather dry but near the timberline 1500 mm have been measured (Hurni 1982). Savannahs below 2000 m are followed by evergreen montane forest with Cedar, Olea europaea, Rapanea simensis and finally Kosso between 2000-3500 m (3300 m in the southwest). Between 2900 and 3700 m ericaceous vegetation takes over, but here Erica arborea is the only tree heather; joined by Hypericum revolutum, as well as Nuxia congesta in the lower elevations (Klötzli 1975; Verfaille 1978; Nievergelt et al. 1998). Above this belt, Festuca macrophylla, Carex erythrorhiza, and Lobelia rhynchopetalum occur, the latter with an inflorescence up to 5m tall. In the highest regions, stands are gradually replaced by Festuca abyssinica and finally by Helichrysum citrispinum scrub up to the summit region.
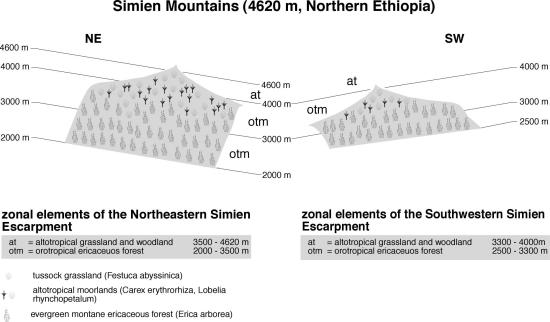
Fig. 10. Vegetation zonation in the Simien Mts.
Maloti-Drakensberg: Mountain vegetation
in southern Africa
Environmental conditions
The Maloti-Drakensberg forms the highest part of the Great Escarpment which bounds the interior plateau of southern Africa. The geological structure is relatively homogenous and characterized by stratified Jurassic flood basalts in the upper portion, which cap the underlying fine-grained Clarens Sandstone (c. 1800-2200 m) and other formations of the Karroo Supergroup. For approximately 200 kilometers the towering scarp of the High Drakensberg constitutes the boundary between the eastern highlands of Lesotho and South Africa. An abrupt and rugged barrier ranging from the foothills at about 1400 m to numerous peaks, buttresses, pinnacles, and cutbacks of the escarpment edge lying roughly between 2800 and 3300 m marks the declivity towards South Africa. Some long spurs (the so-called Little Berg) run out at right angles to the escarpment. The mountain divide consists of two parts: the northern Berg, a north-east-facing portion between Mont aux Sources (3282 m) and Giant's Castle (3314 m), and the southern Berg, a south-east-facing part of the escarpment between Giant's Castle and Wilson's Peak (3310 m). Towards the west, the summit plateau gently slopes down to the foothills and lowlands of Lesotho below 2000 m. Rounded slopes, small cliffs and deeply incised valleys characterize the treeless plateau. The highest peak Thabana Ntlenyana (3482 m) lies about 4 km back from the edge of the escarpment (Hilliard & Burtt 1987; Killick 1990).
Mean annual precipitation in this high mountain area ranges from 1240 mm at the foot of the escarpment (1370 m) to a maximum of about 2000 mm at an altitude of 2300 m. The escarpment edge receives about 1600 mm (rainfall data from the Cathedral Peak area after Killick 1963). Letseng-la-draai (3050 m), located in the rain shadow receives 713 mm, and Mokhotlong Town (2377 m) about 35 km to the west of the escarpment receives only about 575 mm (Killick 1978b). Apart from these steep gradients in rainfall distribution, the humidity conditions are generally characterized by strong seasonal differences. Almost 80 % of the annual precipitation falls between October and March, resulting in generally humid conditions during the growing season (Killick 1978b). Frequent thunderstorms and recurrent periods of fog, which usually do not extent into Lesotho for more than 3 km contribute to summer humidity. Snow falls frequently between April and September and is usually restricted to the summit plateau and nearby slopes of the escarpment. Snow cover seldom lasts for longer periods, in exceptional years it may last for two months. Recurrent droughts force plants to sustain water stress. Generally, the southern Drakensberg is colder and drier than the northern portion. The temperature regime in the upper valley sections near the escarpment is characterized by a long frost period, lasting for approximately 180 days (Grab 1997a). Mean annual temperature is 5.7°C in Letseng-la-draai, and 11.5°C in Mokhotlong Town. Absolute minimum temperature exceeds -20°C at the highest altitudes. Therefore, periglacial conditions with Thufur, needle ice, patterned ground, and soil movements are common at higher elevations (Grab 1994, 1997b, 1998). Prominent differences between the warmer and drier north facing slopes and the colder and moister southerly slopes result from different solar radiation income, especially during winter (Granger & Schulze 1977). Obviously, differences of snow cover duration and soil moisture are ecologically most important and mirrored by a more lush vegetation of the southerly exposures. Most plant species of the Drakensberg are adapted to intense solar radiation, long periods of low temperature and drought, and solifluction due to frost heaving. The frequency of strong winds, especially in spring and summer is considered to be "a powerful factor militating against tree growth" by Hilliard & Burtt (1987).
Phytogeographical
position and altitudinal zonation: Problems of nomenclature
The
vertical sequence of the Maloti Drakensberg has been differentiated
into a montane (approximately 1280-1830 m), subalpine
(1830-2870 m) and alpine belt by Killick (1963, 1990). Other
studies like those of Herbst & Roberts (1974) and Morris et al.
(1993) follow this altitudinal division. White (1983) includes the
"afroalpine" belt as an "archipelago-like region of extreme
floristic impoverishment" within the "afromontane region" On
the other hand, Hilliard & Burtt (1987) reject the possibility to
include the slopes and summits of the Maloti-Drakensberg in an
"afromontane" and "afroalpine" region on the basis that
floristic affinities with adjacent lowlands of southern Africa are
stronger than with the tropical mountains of Tanzania and Kenya. They
argue for the recognition of the area as a distinct phytochorion, the
"South-eastern Mountain Regional Mosaic" instead. In the most
recent overview of the vegetation of southern Africa, Cowling &
Hilton Taylor (1997) include the Drakensberg and Lesotho mountains
together with the mountains of the Cape Folded Belt in an
"afromontane region". In his latest paper Killick (1997)
describes the treeless vegetation of the summit plateau as tundra.
The proposed nomenclature postulates a low-lying laural belt with
supralaural forests up to approximately 2000 m and largely
tree-deficient grass-savannas on the upper mountain slopes.Altitudinal zonation of vegetation
Themeda
triandra grasslands with Trachypogon spicatus, Hyparrhenia
hirta, Harpochloa falx predominantly covers the slopes between
approximately 1600 and 2750 m. Large tussocks of
Merxmuellera macowanii (1830-2550 m) and Festuca
costata (below 2900 m) occur along the streams. Depending on
the kind of fire regime some of these grass slopes up to about 2400 m
contain open Protea savannas, including 3-5 m high trees
of Protea caffra and P. roupelliae with rounded crowns,
and the fire-resistant dwarf species Protea dracomontana.
Locally, 2-4 m high Aloe ferox (=A. candelabrum),
characteristic for drier conditions can be found as a grassland
constituent on steep rocky north-facing slopes in the southern
Drakensberg (Loteni valley) (Hilliard & Burtt 1987, Killick
1990). Small forest groves with the 12-18 m high Yellowwood
(Podocarpus latifolius, up to 1900 m) and Olinia
emarginata (up to 2150 m), often festooned with Usnea
are confined to narrow valleys and southern aspects along stream
gullies. These forests are larger and more numerous on the northern
Berg (Hilliard & Burtt 1987). The forest margins are
characterized by the Rosaceae Leucosidea sericea, the
commonest woody plant on the Drakensberg (up to 2100 m),
Buddleia salviifolia and Bowkeria verticillata,
or by up to 2.5 m high Miscanthus capensis-Cymbopogon validus
grasslands and the Berg bamboo Thamnocalamus tessellatus
(1800-2000 m). The altitudinal distribution of forest
margins roughly extends from the valley floors to the lowermost
basalt cliffs at about 2000 m; some tree clumps persist up to
2400 m (Hilliard & Burtt 1987). Evergreen thickets with the
Thymelaeaceae Passerina drakensbergensis (2100-2450 m on
the northern Drakensberg), the Ericaceae Phillipia evansii
(1800-2300 m), the Compositae Euryops tysonii
(1800-2500 m) and the Cupressaceae Widdringtonia nodiflora
("Subalpine Fynbos" after Killick 1963) are limited
in extent because of recurrent grass fires. Forest in Lesotho only
exists to a limited extent of 1 km2 (Leslie 1991, Low &
Rebelo 1998) with the largest stand in Tsehlanyane (2100 m),
which accommodates a 6 m high Leucosidea sericea forest.The vegetation of the summit plateau of Lesotho between approximately 2850 m and the highest peaks comprise grasslands, dwarf scrub communities, wetland communities along the streams and moister slopes, and open scree and rock communities, colonized by small and scattered turf patches along the basalt outcrops. These vegetation types are best developed from December to the end of March. The grasslands are usually dominated by the Gramineae Merxmuellera disticha, Festuca caprina, Pentaschistis oreodoxa, Harpochloa falx and the Cyperaceae Scirpus falsus in moister places. Subtropical Themeda triandra grasslands are confined to the lower slopes and warmer north-facing aspects below 3000 m. Generally, species of temperate affinity like Festuca caprina are more abundant on the cooler aspects and at higher altitudes. These mixed grasslands are distributed as mosaics, often interspersed by dwarf scrubs and patches of up to 1 m high Merxmuellera drakensbergensis stands at water surplus sites. The evergreen tussock grass is adapted to low temperatures through accumulation of dead and decaying material that acts as an insulator at the base of the plant. The most common dwarf scrubs are Helichrysum trilineatum and Erica dominans, covering considerable areas with low heathlands, especially in the vicinity of Mont Aux Sources. Apparently, this led Killick (196, 1979) to consider Erica-Helichrysum heathland to be the climax community of the uppermost altitudinal belt. The cushion-forming succulent Euphorbia clavarioides is limited to steep rocky and mostly north-facing slopes (Hilliard & Burtt 1987). Furthermore, the two karroid invader species Chrysocoma ciliata and Pentzia cooperi (Morris et al. 1993) together with Artemisia afra dominate the dwarf scrub communities between Sani Pass and Mokhotlong. Wetlands and flat marshy grasslands in the riverheads contrast to the surrounding vegetation. The low-growing wetland vegetation contains a mixture of Gramineae (Agrostis sp., Poa annua) and Cyperaceae (Scirpus falsus, S. ficinioides, Isolepis fluitans and Schoenoxiphium filiforme). Pools and silt patches, often waterlogged in summer form a habitat for aquatic communities with Limosella inflata, Aponogeton ranunculiflorus and Crassula inanis surrounded by Merxmuellera drakensbergensis tussocks and hygrophilous forbs like Ranunculus meyeri, Kniphofia caulescens, Moraea alticola frequently accompanied by Trifolium burchellianum (van Zinderen Bakker 1955, Killick 1978c, Backéus & Grab 1995, Schwabe 1995). Thufur are often colonized by Athrixia fontana.
Acknowledgements
The financial support of DFG (German Science Foundation) for large parts of the fieldwork of this study is gratefully acknowledged. The author wishes to thank Karsten Wesche and Wolfgang Nuesser for their contributions on the vegetation of the Atlas and the Drakensberg, and the regeneration of Erica dominated forests on the Afrotropical Mountains.
References
Allaud, C. & R. Jeannel. 1914. Le Mt. Kenya. Rev. Gen. Sci. 25, 639-644.
Backéus, I & S. Grab. 1995. Mires in Lesotho. Gunneria 70, 243-250.
Beck, E. 1994. Cold tolerance in tropical alpine plants. Rundel, P.W., A.P. Smith & F.C. Meinzer. (Eds.). Tropical alpine environments. Cambridge University Press. Cambridge. 77-110.
Beck, E., H. Rehder & J.O. Kokwaro. 1990. Classification and mapping of the vegetation of the alpine zone of Mount Kenya (Kenya). - Geographica Bernensia African Studies Series A8. Mt. Kenya Area, 41-46.
Berger, P. 1989. Rainfall and Agroclimatology of the Laikipia Plateau, Kenya. Geographica Bernensiaca - African Studies Series A7, 1-102, 3 maps.
Biedinger, N. 1995. Ökologie und Diversität der Epiphyten-Vegetation in zentralafrikanischen Wäldern (Ruanda, Zaire). - PhD thesis. Mathematisch-Naturwissenschaftliche Fakultät. Rheinische Friedrich-Wilhelms-Universität. Bonn, 226.
Bussmann R.W. 1994. The forests of Mount Kenya (Kenya) - Vegetation, Ecology, Destruction and Management of a Tropical Mountain Forest Ecosystem. (Dissertation Universität Bayreuth, Vol. 1-3).
Bussmann, R.W. 1997. The forest vegetation of the Harenna Escarpment (Bale Province, Ethiopia) - syntaxonomy and phytogeographical affinities. Phytocoenologia 27, 1-23.
Bussmann, R.W. 2001. Succession and regeneration patterns of East African mountain forests. Systematics and Geography of Plants 71, 959-974.
Bussmann R.W. & E. Beck 1995a. The forests of Mount Kenya (Kenya), a phytosociological synopsis. Phytocoenologia 25 (5), 467-560.
Bussmann R.W. & E. Beck. 1995b. Regeneration- and cyclic processes in the Ocotea-Forests (Ocotea usambarensis Engl.) of Mt. Kenya. Verh. Ges. Ökologie 24, 35-38.
Bussmann R.W. & E. Beck. 1995c. Regeneration- and succession processes in the Cedar-Forests (Juniperion procerae) of Mt. Kenya. Ecotropica 1, 79-84.
Bussmann R.W. & S. Lange. 1999. Succession and regeneration patterns of East African mountain forests. In: BITÖK (Ed.) Vegetationsökologische, ethnobotanische und -faunistische Beiträge über Äquatorial-Afrika. Bayreuther Forum Ökologie 64, 1-16.
Bussmann R.W. & E. Beck. 1999. Vegetation units of Mount Kenya Forest Reserve - Comment. In: BITÖK (Ed.) Vegetationsökologische, ethnobotanische und -faunistische Beiträge über Äquatorial-Afrika. Bayreuther Forum Ökologie 64, 17-28.
Bytebier, B. & R.W. Bussmann. 2000 / printed 2002. Vegetation of Mount Nyiru (Samburu District, Kenya), A checklist and syntaxonomical survey. J. E. Afr. Nat. Hist. 89, 45-71.
Cable, S. & M. Cheek, M. 1998. The plants of Mt. Cameroon. Royal Botanical Gardens. Kew. 198 pp.
Cowling, R.M. & C. Hilton-Taylor 1997. Phytogeography, Flora and Endemism. Cowling, R.M., D.M. Richardson & S.M. Pierce (Eds.). Vegetation of Southern Africa. Cambridge, University Press, 43-61.
Decurtins, S. 1985. Hydrogeographical investigations on Mt. Kenya and its w and nw slopes. Laikipia Reports 5, 1-14.
Decurtins, S. 1992. Hydrographical investigations in the Mount Kenya subcatchment of the Ewaso Ngiro River. Geographica Bernensiaca - African Studies Series A10, 1-151.
Engler, A. 1895. Die Pflanzenwelt Ostafrikas und der NachbargebieteBerlin. pp.
Fischer, E. 1996. Die Vegetation des Parc National de Kahuzi - Biega, Sud Kivu, Zaire. Franz Steiner Verlag. Stuttgart. 240 pp.
Grab, S. 1994. Thufur in the Mohlesi valley, Lesotho, Southern Africa. Permafrost and Periglacial Processes 5, 111-118.
Grab, S. 1997a. Analysis and characteristics of high altitude air temperature data from northern Lesotho. Implications for cryogeomorphic occurrences. Geooekoplus 4, 49-60.
Grab, S. 1997b. Thermal Regime for a Thufa Apex and its Adjoining Depression, Mashai Valley, Lesotho. Permafrost and Periglacial Processes 8, 437-445.
Grab, S. 1998. Non-Sorted Patterned Ground in the High Drakensberg, Southern Africa, Some New Data. The Geographical Journal 164, 19-31.
Granger, J.E. & R.E. Schulze 1977. Incoming Solar Radiation Patterns and Vegetation Response, Examples from the Natal Drakensberg. Vegetatio 35, 47-54.
Hall, J.B. 1973. Vegetational zones of the southern slopes of Mount Cameroon. Vegetatio 27, 19-69.
Hauman, L. 1955. La "region afroapline" en phytogeographie centro africaine. Webbia XI, 467-489.
Hedberg, O. 1951. Vegetation belts of the East -African mountains. Svensk Botanisk Tidskrift 45, 141-196.
Hedberg, O. 1955. Altitudinal zonation of the vegetation on the east African Mountains. Proceedings of the Linnean Society of London (Botany) 165, 134-136.
Hedberg, O. 1964. Features of afro-alpine plant ecology. Acta Phytogeographica Suecica 49, 1-144.
Heinrichs, T. 2001. Geologie - Ostafrika. Gebrüder Borntraeger. Berlin, Stuttgart. 231 pp.
Hemp, A. 2001. Ecology of the pteridophytes on the southern slopes of Mt. Kilimanjaro. Part II, Habitat selection. Plant Biology 3, 493-523.
Hemp, A. 2002. Ecology of the pteridophytes on the southern slopes of Mt. Kilimanjaro. I. Altitudinal distribution. Plant Ecology 159, 211-239.
Hemp, A. & E. Beck. 2001. Erica excelsa as a fire-tolerating component of Mt. Kilimanjaro's forests. Phytocoenologia 31, 449-475.
Herbst, S.N. & B.R. Roberts 1974. The Alpine Vegetation of the Lesotho Drakensberg, A Study in Quantitative Floristics at Oxbow. Journal of South African Botany 40, 257-267.
Hilliard, O.M. & B.L. Burtt 1987. The Botany of the Southern Drakensberg. Cape Town. Annals of the Kirstenbosch Botanic Gardens 15.
Hurni, H. 1982. Klima und Dynamik der Höhenstufung von der letzten Kaltzeit bis zur Gegenwart. Geographica Bernensia G 13.
Hutchins, D.E. 1909. Report on the forests of British East Africa. London, Darling & Son.
Jackson, J.K. 1956. The vegetation of the Imatong Mts., Sudan. J. Ecology 44, 341-374.
Karlowski, U. 1995. Sekundäre Sukzession im afromontanen Nebelwald, Dynamik, Mechanismen und Schutz der Biodiversität in zwei Habitaten des Berggorillas (Mgahinga Gorilla- und Bwindi Impenetrable-Nationalpark) und im Echuya Forest, Uganda. - PhD. Mathematisch Naturwissenschaftliche Fakultät. Rheinische Friedrich Wilhelms Universität. Bonn, 178.
Killick, D.J.B. 1963. An Account of the Plant Ecology of the Cathedral Peak Area of the Natal Drakensberg. Pretoria. Memoirs of the Botanical Survey of South Africa 34.
Killick, D.J.B. 1978a. The Afro-Alpine Region. In: Werger, M.J.A. (Ed.). Biogeography and Ecology of Southern Africa. The Hague, Junk. Monographiae Biologicae 31, 515-560.
Killick, D.J.B. 1978b. Further Data on the Climate of the Alpine Vegetation Belt of Eastern Lesotho. Bothalia 12, 567-572.
Killick, D.J.B. 1978c. Notes on the Vegetation of the Sani Pass Area of the Southern Drakensberg. Bothalia 12, 537-542.
Killick, D.J.B. 1979. African Mountain Heathlands. In: Specht, R.L. (Ed.). Heathlands and Related Shrublands (Ecosystems of the World 9A). Amsterdam, Elsevier, 97-116.
Killick, D.J.B. 1990. A Field Guide to the Flora of the Natal Drakensberg. Johannesburg, Jonathan Ball and Ad Donker.
Killick, D.J.B. 1997. Alpine Tundra of Southern Africa. In: Wielgolaski, F.E. (Ed.). Polar and Alpine Tundra (Ecosystems of the World 3). Amsterdam, Elsevier., 199-209.
Klötzli, F. 1975. Zur Waldfähigkeit der Gebirgssteppen Hoch-Semiens. Beitr. naturk. Forsch. Südw. Dtl. Band 34, 131-147.
Knapp, R. 1973. Die Vegetation von Afrika unter Berücksichtigung von Umwelt, Entwicklung, Wirtschaft, Agrar- und Forstgeographie. G. Fischer. Jena. 626 pp.
Knox, E.B. 1993. The conservation status of the giant senecios and giant lobelias in Eastern Africa. Opera Botanica 121, 195-216.
Leibundgut, C. et. al. 1986. Hydrogeographical Map of Mount Kenya Area. Geographica Bernensiaca - African Studies Series A3, 1-70, map.
Leslie, A.D. 1991. Indigenous Forest and Woodland in the Kingdom of Lesotho. Suid-Afrikaanse Bosboutydskrif 158, 97-103.
Negatu, L. & M. Tadesse. 1989. An ecological study of the vegetation of the Harenna forest, Bale, Ethiopia. Sinet 12, 63-93.
Low, B.A. & A.G. Rebelo. 1998. Vegetation of South Africa, Lesotho and Swaziland. Pretoria, Department of Environmental Affairs and Tourism.
Mabberley, D.J. 1975. Notes on the vegetation of the Cherangani Hills, N.W. Kenya. Journal of the East African Natural History Society and National Museum 15, 1-11.
Maitland, T.D. 1932. The grassland vegetation of the Cameroon Mountain. Bulletin of miscellaneous information, Royal Botanical Gardens, Kew 9, 417-435.
Gashaw, M. & M. Fetene. 1996. Plant communities of the afroalpine vegetation of Sanetti Plateau, Bale Mountains, Ethiopia. Sinet 19, 65-86.
Miehe, S. & G. Miehe. 1994. Ericaceous forests and heathlands in the Bale Mountains of South Ethiopia - Ecology and man's impact. Stiftung Walderhaltung in Afrika. Hamburg. 206 pp.
Miehe, S. 1988. Vegetation ecology of the Jebel Marra Massif in the Semiarid Sudan. J. Cramer. Berlin, Stuttgart. 171 pp.
Morris, C.D., N.M. Tainton & S. Boleme. 1993. Classification of the Eastern Alpine Vegetation of Lesotho. African Journal of Range and Forage Science 10, 47-53.
Ndam, N., J. Healey, M. Cheek & P. Fraser. 2001. Plant recovery on the 1922 and 1959 lava flows on Mount Cameroon, Cameroon. Systematics and Geography of Plants 71, 1023-1032.
Neville, G. & K. Wesche. (in press). A land-cover and vegetation map of the eastern slopes of Mt. Elgon. - Mountain Research and Development.
Nievergelt, B., T. Good & R. Güttinger. 1998. A survey on the flora and fauna of the Simen Mountains National Park. Walia (special issue), 1-109.
Oliver, E.G.H. 1987. Studies in the Ericoideae (Ericaceae). VII. The placing of the genus Philippia into synonomy under Erica, the southern Africa species. South African Journal of Botany 53, 455-458.
Osmaston, H. & G. Kaser. 2001. Rwenzori Mountains National Park, Uganda. Glaciers and Glaciations. Gärtner, E. & Georger, C. Glasgow, Bartholomew.
Osmaston, H. 1996. Glaciations, landscape and ecology. Osmaston, H., C. Basalirwa & J. Nyakaany (Eds.). The Rwenzori Mountains National Park, Uganda. Makarere University. Kampala. 49-65.
Petters, S.W. 1991. Regional geology of Africa. Springer. Berlin, Heidelberg. 722 pp.
Rauh, W. 1952. Vegetationsstudien im Hohen Atlas und dessen Vorland. Sitzungberichte der Heidelberger Akademie der Wissenschaften - Mathematisch naturwissenschaftliche Klasse 1-118.
Rehder, H., E. Beck, J.O. Kokwaro & R. Scheibe. 1981. Vegetation Analysis of the upper Teleki Valley (Mount Kenya) and adjactent Areas. J. E. Afr. Nat. Hist. Soc. Nat. Mus. 171, 1-8.
Rehder, H., E. Beck & J.O. Kokwaro. 1988. The afroalpine plant communities of Mt. Kenya (Kenya). Phytocoenologia 16, 433-463.
Richards, P.W. 1963. Ecological notes on west African vegetation III. The upland forests of Cameroon Mountain. Journal of Ecology 51, 529-554.
Schlüter, T. 1997. Geology of East Africa. Borntraeger. Berlin. 484 pp.
Schmitt, K. 1991. The Vegetation of the Aberdare National Park Kenya. Wagner. Innsbruck. 259 pp.
Schmitt, K. 1992. On the afroalpine vegetation of the Ruwenzori Mountains, Uganda. - Phytocoenologia 21, 313-322.
Schwabe, C.A. 1995. Alpine Mires of the Eastern Highlands of Lesotho. Cowan, G.I. (Ed.), Wetlands of South Africa. Pretoria, Department of Environmental Affairs and Tourism.
Snowden, J.D. 1933. A study of the altitudinal zonation in South Kigezi and on Mounts Muhavura and Mgahinga, Uganda. Journal of Ecology 21, 7-27.
Tamrat, B. 1993. Vegetation ecology of remnant Afromontane forests on the Central Plateau of Shewa, Ethiopia. Opulus Press AB. Uppsala. 64 pp.
van Heist, M. 1994a. Accompanying report with the Land unit map of Mount Elgon National Park. Mount Elgon Conservation and Development Project. Kampala. 83 pp.
van Heist, M. 1994b. Land unit map of Mount Elgon. Kampala, Mt. Elgon Conservation and Development Project.
van Heist, M. 1999. Land unit map the Ruwenzori Mts. National Park. Fort Portal, WWF-RMCDP.
van Zinderen Bakker, E.M. 1955. A Preliminary Survey of the Peat Bogs of the Alpine Belt of Northern Basutoland. Acta Geographica 14, 413-422.
Verfaille, M. 1978. The Ericaceous Belt of the Semien Mountains National Park, Ethiopia. - Biol. Jb. Dodonea 46, 210-223.
Weinert, E. 1983. Vegetation zones in Bale Mountains near Goba, S. Ethiopia. - Wissenschaftliche Zeitschrift der Martin Luther Universität Halle Wittenberg 1983, 41-67.
Weischet, W. & W. Endlicher. 2000. Regionale Klimatologie. Teil 2. Die Alte Welt. Teubner. Stuttgart, Leipzig. 625 pp.
Wesche, K. 2002. The high-altitude environment of Mt. Elgon (Uganda/Kenya) - Climate, vegetation and the impact of fire. - Ecotropical Monographs 2, 1-253.
Speck H. 1986. Soils of the Mount Kenya Area, their formation, ecological, andagricultural significance. Winiger, M. (Ed.) Mt. Kenya Area. Contributions to Ecology and Socio-Economy. African Studies Series A1, 47-72.
White, F. 1983. The Vegetation of Africa. Paris, Unesco.
White, F. 1950. The forests of Mount Kenya. Forest Society Journal Oxford 3(5), 32-38.
48