


 Email this Article
Email this Article Print
Print Download (PDF)
Download (PDF)
Resumen 
Determinamos la composición faunística de quirópteros en la vertiente norte del Parque Nacional Henri Pittier, localizado en los 10° 21' 33" y 10° 31 '17" de latitud norte y los 67° 33' 14" y 67° 39' 38" de longitud oeste. Aunque no existen políticas claras de conservación para los cultivos tradicionales de cacao dentro del parque, estos constituyen importantes reservorios de biodiversidad. Se muestrearon las comunidades de quirópteros de una plantación de cacao y un bosque seco, con un esfuerzo de captura de 1.008 horas - malla, encontrándose una proporción de 84% Phyllostomidae, 7% Vespertilionidae, 5% Desmodontidae, 2% Mormoopidae y 2% Emballonuridae respectivamente. Las especies más abundantes en ambos ambientes (Carollia perspicillata y Artibeus jamaicensis) se alimentan de frutos asociados a especies vegetales sucesionales que conforman los bosques que protegen al cacao o están presentes en el bosque seco donde abundan plantas de Cecropia sp. Para comparar diversidad en ambos ambientes se calcularon valores de alfa, beta y gamma. La diversidad beta encontrada nos permite afirmar que los murciélagos aprovechan la estructura florística del cacaotal que se encuentran en algunos fragmentos del parque como proveedor de recursos ecológicos a numerosas especies las cuales son beneficiarias directas del microhabitats que allí encuentran.
Palabras claves: quirópteros, recursos ecológicos, alfa, beta y gamma diversidad.
Abstract
We determine the composition of chiropteran's fauna in the north slope of the Henri Pitter's National Park, located in the 10° 21' 33" and 10° 31 '17" of north latitude and the 67° 33' 14" and 67° 39' 38" of west longitude. Although there are not clear politics of conservation for the traditional cultives of cocoa inside the park, they constitute important biodiversity reservoirs. Sampling the chiropteran communities of cocoa plantation and dry forest, with a capture effort of 1.008 hours - mesh, having found a proportion of 84% Phyllostomidae, 7% Vespertilionidae, 5% Desmodontidae, 2% Mormoopidae and 2% Emballonuridae respectively. The most abundant species in both regions (Carollia perspicillata and Artibeus jamaicensis) they feed fruits associated to species succession plants which conform the forests that protect the cocoa or that are present in the dry forest where plants of Cecropia sp are plentiful. In order to compare the diversity in both regions the alpha, beta and gamma values were calculated. The beta opposing diversity allows us to affirm that the bats take advantage of the floristic structure of the cocoa plantation that are in some areas of the park as supplier of ecological resources to numerous species which are direct beneficiaries of the microhabitats found.
Key words: chiropteran, ecological resources, alpha, beta, and gamma diversity.
Introducción
Aunque no existen políticas claras de conservación para los cultivos tradicionales de cacao dentro de los parque nacionales que los poseen aunque estos constituyen importantes reservorios de biodiversidad (Moreno, C 2000). El vocablo biodiversidad en el ámbito científico es difícil de estandarizar. Incluso hay detractores del término que argumentan que es tal la magnitud conceptual de este neologismo que expresa todo y, al mismo tiempo, nada. Algo hay de razón en ello, pero sólo algo. Para Hurlbert (1978), la diversidad es un 'non-concept' Otros en cambio, afirman que no es fácil definirla, debido a que la diversidad en un contexto ecológico, engloba dos componentes: variedad y abundancia relativa de especies (Magurran 1988).Cuando nos referimos a biodiversidad en un contexto conservacionista, estamos hablando de diversidad de especies, de variación intraespecífica e intrapoblacional, y en última instancia de variación genética, que no por estar enmascarada a veces por fenómenos de dominancia y expuesta a la desaparición deja de ser importante (Ezcurra 1990). La diversidad biológica puede conceptualizarse generalmente como la variedad total de la vida o la propiedad que caracteriza a los seres vivos de estar formados por diversas entidades. En el ámbito ecológico Halffer & Ezcurra (1992). definen la biodiversidad en dos categorías: la diversidad alfa y la diversidad beta. La primera representa una función de la cantidad de especies presentes en un mismo habitad y la segunda comprende una medida del grado de participación del ambiente en parches o mosaicos biológicos, es decir, mide la continuidad de hábitats diferentes en el espacio. Solbrig (1991) se refiere a esta última como las diferencias entre comunidades que mide la tasa de cambio entre hábitats. Según Moreno (2000), esta proporción puede evaluarse en base a índices o coeficientes de similitud, disimilitud de distancia entre las muestras a partir de datos cualitativos o cuantitativos. En un sentido estricto, la diversidad, un concepto derivado de la teoría de sistemas, es simplemente una medida de la heterogeneidad del mismo. En el caso de los sistemas biológicos, la diversidad se refiere a la heterogeneidad biológica, es decir, a la cantidad y proporción de los diferentes elementos biológicos que contenga el sistema. La estimación de la biodiversidad depende, entre otras cosas, de la escala a la cual se defina el problema (Halffer & Ezcurra 1992).
Venezuela se ubica entre los primeros diez países con mayor diversidad biológica del planeta y el sexto en América. Esta condición se debe, en gran parte, a la convergencia de cuatro importantes regiones biogeográficas, como son la Amazónica, la Andina, la Caribeña y la Guayanesa, que le confieren una amplia diversidad de biomas representados en las diferentes provincias naturales. Esta especial bondad natural y geográfica la ubica entre los países megadiversos del planeta, ventaja comparativa poco evidenciada y valorada como potencial económico estratégico(Aguilera et al. 2002). En relación con las plantas se ha estimado que las dicotiledóneas (10.110 especies) constituyen más de 60% de las especies vegetales existentes en el país. Dentro de las plantas del nuevo mundo encontramos el cacao (Theobroma cacao), este cultivo esta señalado para los valles de las costas aragüeñas desde 1610 y desde 1616 constituye una siembra sistemática en la localidad de Choroní.(Varela 1999). Su producción declina fuertemente una vez que estas áreas son declaradas parque nacional en el año 1937, manteniéndose aun como un cultivo tradicional con poca intervención antropogénica y constituye el sustento económico de gran parte de la población de la zona.
El cultivo de cacao constituye un sistema de importancia ya que aporta una cantidad significativa de materia orgánica por lo tanto beneficia a la microflora y la fauna del suelo. Además de esto, el cacaotal es un reservorio de invertebrados y vertebrados que de alguna manera interaccionan para mantener la dinámica poblacional de estos agroecosistemas. La escasa literatura donde se señalan los aportes de las plantaciones de cacao al mantenimiento de la biodiversidad y los niveles de conexión que se establecen entre los bosques que por lo general rodean estos cultivos y la fauna que allí se alberga fue uno de los motivos para la realización de este trabajo. Se hace énfasis en el estudio de las poblaciones de quirópteros presentes en este cultivo y su relación. El conocimiento de la biología y ecología de los vertebrados como parte de los recursos naturales que habitan en un área determinada constituyen información fundamental a la hora de tomar decisiones para reglamentar su uso y manejo. Los ambientes naturales presentan alta disponibilidad de recursos y se alteran con la introducción del cultivo de cacao. Aunque en las costas aragüeñas el cultivo del cacao por tener un manejo tradicional basado en un bajo nivel tecnológico y estar rodeado de la cordillera montañosa que conforma el Parque nacional Henri Pittier, tal vez ofrezca un ambiente muy particular con cierto grado de equilibrio ecológico en las plantaciones que hacen de ellas un lugar que de algún modo provee recursos o sirve de refugio para muchas especies de fauna (Ortiz 1993). Este estudio esta centrado en medir la diversidad de los murciélagos y sus relaciones con el ambiente, puesto que estos constituyen un grupo animal muy importante dentro del complejo funcionamiento de los ecosistemas y mantienen bajas algunas poblaciones de insectos, son responsables de la polinización de muchas especies vegetales y de la dispersión de muchas semillas (Fleming et al. 1972). No obstante, la estimación de dicha diversidad y abundancia de algunas especies estará limitada por el método de captura a utilizar (redes de neblina). Sin embargo, el sesgo podría no ser muy grande en el inventario de especies (Sokal & Rohlf 2000), con potencial falla de detección de algunos emballonúridos y vespertilionidos presentes debajo del dosel dentro del cacaotal ya que probablemente los insectívoros de vuelo libre más rápidos utilicen estratos superiores para su alimentación.
Materiales y Métodos
Área de estudio
El Parque Nacional Henry Pittier está ubicado en la parte centro- norte de Venezuela, sobre la rama litoral de la cadena montañosa conocida como Cordillera de la Costa o Caribe, cuyo eje principal o fila maestra, lo divide en una vertiente hacia el norte y otra hacia el sur. Sus coordenadas extremas son al norte, 10º 32' 40" de latitud norte, en Punta Tuja sobre el Mar Caribe; al sur, 10º 14' 25" de latitud norte, en el extremo sureño del cerro El Mácaro; al oeste, 67º 52' 54" de longitud oeste, en el Pico Jengibre sobre la fila maestra y al este, 67º 24' 36" de longitud oeste, en el Pico Cogollal, también en la fila maestra, abarcando una superficie total de 107.633 hectáreas (Fernández-Badillo 2000).
.
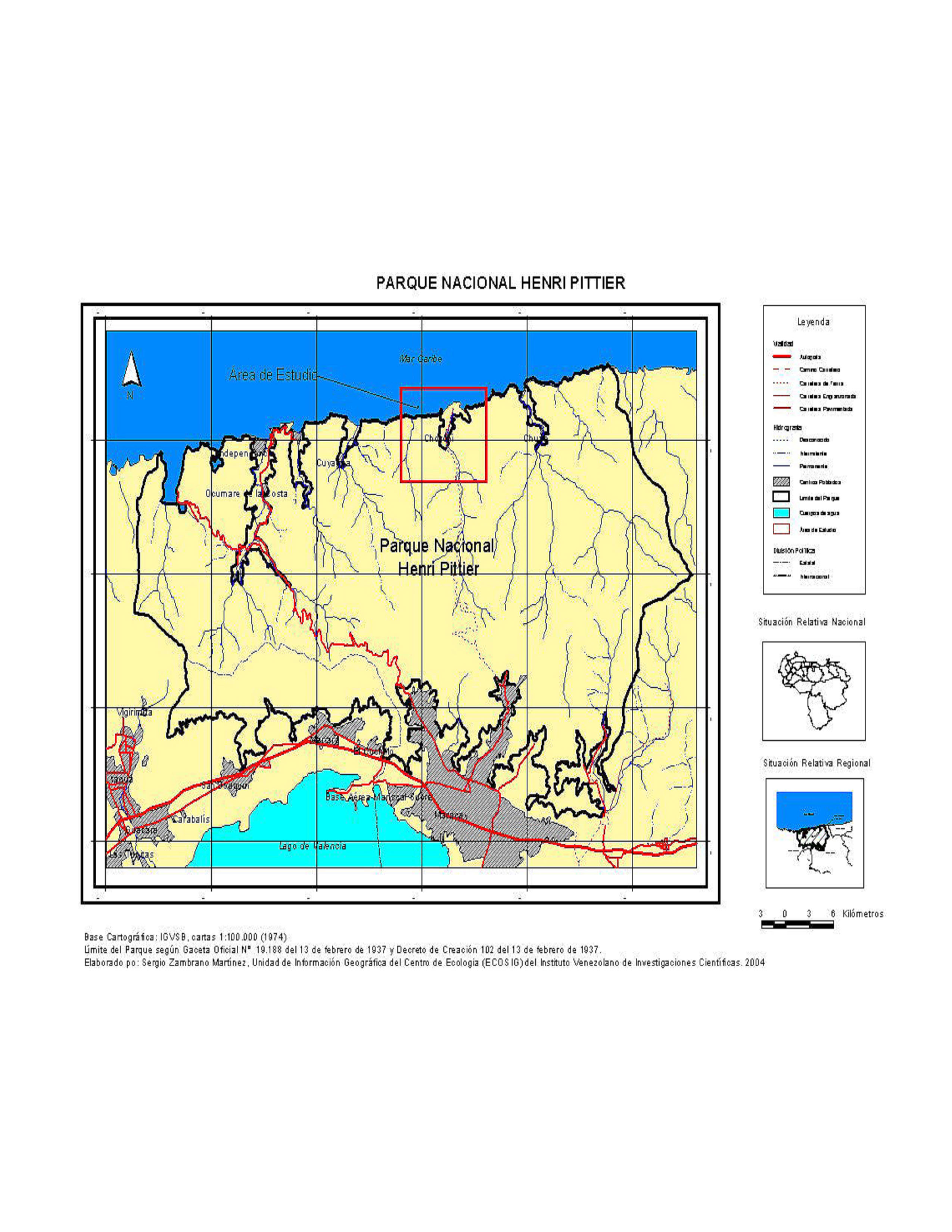
La Parroquia de Choroní tiene un 97% de la superficie bajo la jurisdicción del Parque Nacional Henri Pittier, localizado exactamente entre las coordenadas geográficas de 10° 21' 33" y 10° 31 '17" de latitud norte y los 67° 33' 14" y 67° 39' 38" de longitud oeste, el otro 3% corresponde al valle costanero donde se encuentran los centros poblados de Choroní y Puerto Colombia y alcanza una superficie de 13.800 hectáreas (Manamá & Rivero 1994) es una zona de vida boscosa, decidua, semiárida, tropófila, cálida, de tierras bajas, ya que crece desde unos 200 a 250 m hasta unos 450 a 500 m de altitud (Holdridge 1978). Los Bosques secos de Choroní, se caracterizan por ser más áridos que los bosque de la vertiente sur y están muy relacionados con los cardonales espinares de la franja costera vecina. Un 80% o más de sus especies arbóreas pierden totalmente su follaje durante la época de sequía, presentan tres estratos arbóreos, uno emergente que sobresale sobre las copas hasta una altura de unos 15 m o más, luego un estrato superior y de copas continuas y regulares, al menos durante la época de las lluvias, entre los 10 y 12 m de altura y otro inferior y discontinuo con 6 a 8 m de altura. La fuente de agua proviene de la zona con altitud entre 900 y 2200 m.s.n.m en donde alcanza promedios superiores a los 1800 mm de precipitación anual (Anzoategui 1994). Por el contrario la vertiente norte del parque se caracteriza por ser presentar un bosque seco, deciduo, semiárido, tropófilo, cálido, de tierras bajas, ya que crece desde unos 200 a 250 m hasta unos 450 a 500 m de altitud. Son más secos y áridos que los bosques de la vertiente sur y están muy relacionados con los cardonales espinares de la franja costera vecina. Son abundantes los cactus columnares, entre ellos cardón azul (Cereus hexagonus), cardón viejito (Pilosocereus moritzianus), yaurero (Subpilocereus russelianus) y el cardón de dato (Stenocereus griseus). Luego hay un estrato arbustivo entre 2 a 5 m de altura, donde se encuentran entre otros zapatico (Rauwolfia tetraphylla), basura prieta (Cordia curassavica), pitajaya (Acanthocereus tetragonus), olivo (Capparis indica), carcanapire (Croton spp), cruceto (Randia dioica), zorrocloco (Morisionia americana), cordoncillo (Piper sp).
Materiales
:Para la captura de murciélagos se usaron mallas de neblina CH12: 5/2 38 mm de abertura, 2,6 m de altura por 12 m de ancho, de nylon con 4 bolsillos marca Avinet, colocadas a ras de suelo, en lugares seleccionados dentro de los ambiente objeto de estudio, usando como criterio luego de una previa observación, que fueran los sitios donde existiese la posibilidad de mayor movimiento por parte de los murciélagos phyllostomidos. Se colocaron 2 redes de neblina, las cuales se abrieron durante 3 noches al mes, durante 12 meses, abiertas desde las 18:30 h hasta las 21:30 h y desde las 2:30 h hasta las 6:30 h, para un total de 7 horas por noche, respectivamente durante 3 días consecutivos al mes correspondientes a los periodos de luna nueva con el fin de garantizar la baja luminosidad y tomando en cuenta el éxito de captura las mismas eran revisadas continuamente en el transcurso de la colecta (Wilig 1986), para un total de 36 noches desde diciembre 2001 hasta diciembre 2002 (a excepción de abril 2002, que no se muestreo debido a los problemas políticos que se presentaron en el país), lo cual representa un esfuerzo de captura de 12.096 metros lineales /malla/ hora. Los murciélagos capturados en las mallas fueron sacrificados, posteriormente fijados con formol al 10% y conservados en etanol al 70%. Para el análisis de los datos de bosque ribereño, se utilizó la información generada por Rojas (1994), quien presenta un inventario detallado de especies de murciélagos en un bosque ribereño de la vertiente norte del Parque Nacional Henri Pittier. Su estudio fue realizado muestreando durante un año (desde Agosto de 1993 hasta Agosto de 1994) en la Trilla, para lo cual uso 3 mallas de neblina de 12 metros, las cuales abrió tres noches al mes, cada dos meses, durante un año, para un total de 24 h por mes durante 6 meses, lo que hace un total de 144 horas-malla y 13.478,4 m2 .malla. h. Los datos fueron tomados en base a lo que señalan Moreno & Halffer (2001).con respecto al mínimo esfuerzo requerido para hacer una proyección acerca de la quirópterofauna presente en un área determinada, quienes afirman que es requerido un mínimo de 5 a 18 noches de muestreo para registrar un 90% de la fauna.
Para este estudio se tomaron en cuenta los siguientes datos: localidad, fecha de captura, sexo y condición reproductiva. Los mismos fueron identificados usando la clave taxonómica de Linares (1998) y la clave de mamíferos de Venezuela de Fernández- Badillo et al. (1988) y se verificó la identidad de especimenes preservados en el Museo de la Estación Biológica "Rancho Grande". Para los cálculos estadísticos de los Índices de diversidad se uso el manual de Moreno (2000).
Resultados y Discusión
Los valores de diversidad-alpha de las comunidades de murciélagos nos indican el estado de variación que especies mantienen en poblaciones viables sobre si mismas dentro del hábitat, las mismas puedes estar debidas a elementos transitorios o las llamadas especies turistas (Gaston & Spicer 1988) que pueden llegar desde los alrededores en busca de recursos alimentarios. El arribo de especies desde un determinado hábitat, donde estos no pueden mantenerse a sí mismos, hasta un hábitat adyacente más apropiado, este factor muchas veces estas especies incrementan erróneamente la diversidad -beta de algún hábitat, sin embargo, la probabilidad que esto suceda es muy baja y las especies turistas o transitorias dependen de la movilidad de grupo, de la heterogeneidad espacial y del tamaño y las relaciones topológicas del hábitat (Moreno & Halffer 2001).
La fauna de quirópteros de la vertiente Norte del Parque Nacional Henri Pittier esta compuesta por 34 especies de quirópteros para el cacaotal, 25 para el bosque seco y 25 reportadas por Rojas (1994) para el bosque ribereño. Encontramos hay un decrecimiento en el número de especies capturadas justo los meses que coinciden con la limpia del bosque de cacao. Esta disminución sin embargo no es indicativa de que disminuya la riqueza de especies, puesto que un número alto de especimenes capturados no refleja necesariamente un aumento en la diversidad (Ortiz 1993). Las modificaciones anuales a las que estos bosques de cacao están sujetos, probablemente no inciden sobre su estructura florística y hace que la dinámica de estas plantaciones sea mas plástica que la de otros ecosistemas para tolerar la fragmentación del paisaje por el crecimiento controlado y continuo (replantación) de sus plantas. Es necesario entender la dinámica de los requerimientos de hábitat temporal - espacial de las especies para entender e interpretar los patrones de composición de especies, abundancia, riqueza, por ello hay que distinguir entre las variaciones que son verdaderamente estocásticas y aquellas que son especificas y cuya complejidad causa y efecto no han sido identificadas, aunque se sabe que las variaciones en una comunidad rara vez ocurren aleatoriamente, muchas variables podrían ser deterministicas pero no deben ser interpretadas hacia un solo extremo estos bosques sufren anualmente practicas culturales), asociadas al manejo del cultivo del cacao, por lo cual las variaciones en la composición florística año tras año no determinan la diversidad de vertebrados (Parra 2004)
Sin embargo, Fox & Fox (2000) señalan que la diversidad de hábitats constituye un buen indicador de la riqueza de especies, el incremento en los niveles de distribución por aumento de superficie hará decrecer la riqueza de especie. Obsérvense los resultados obtenidos en cuanto a la proporción de quirópteros asociada a cada hábitat que fue muestreado, demostrándose que el reemplazo de los hábitats prístinos por sucesiones secundarias no necesariamente indicaran el reemplazo o el reestablecimiento de las especies que un momento estuvieron asociadas al bosque original, en caso de hacer comparaciones con muestreos anteriores en la misma área, información que es desconocida.
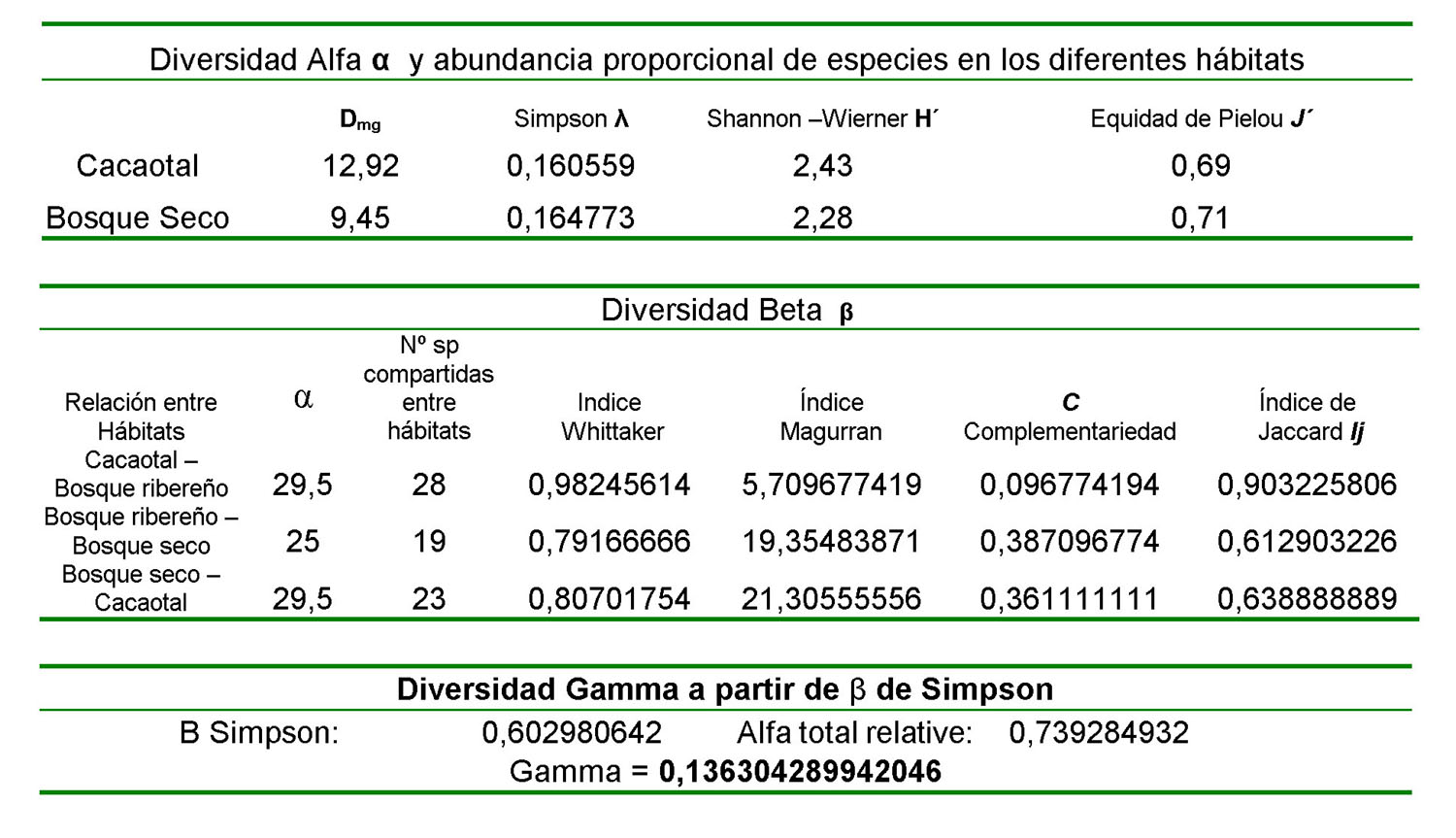
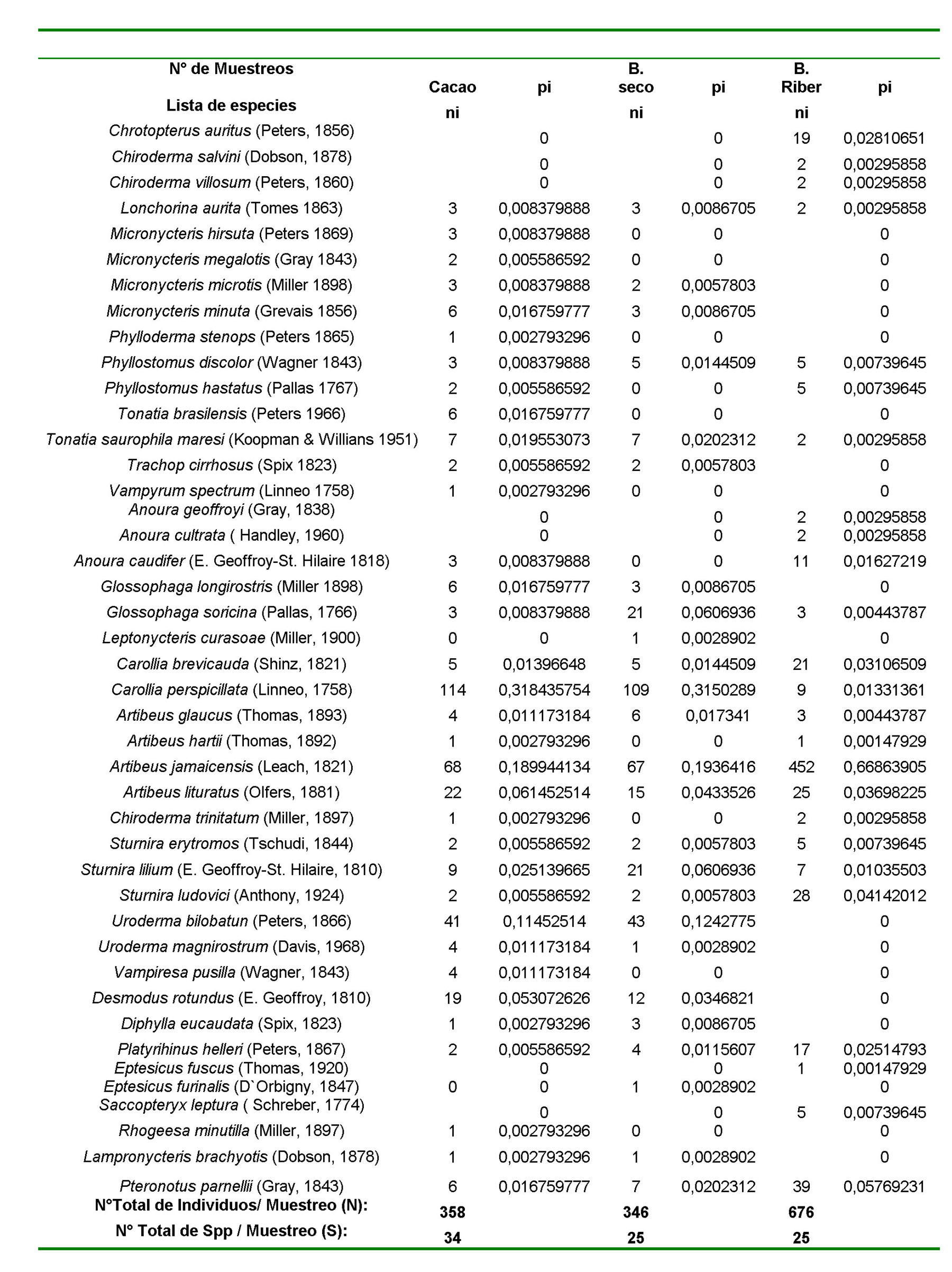
Se capturados 704 murciélagos (358 para el bosque de cacao, 346 para el bosque seco) durante un año de muestreo y se toman los datos reportados por Rojas (1994) para el bosque ribereño, un total de 676 murciélagos, todos estos repartidos en 5 Familias lo cual representa el 28% de los murciélagos reportados para el país. En total de estos el 84% son de la familia Phyllostomidae, el 7% Vespertilionidae, un 5% Desmodidae, 2% Mormoopidae y 2% Emballonuridae pertenecen a esas familias respectivamente. Las especies más abundantes para el bosque de cacao son Carollia perspicillata, Artibeus jamaicensis, Uroderma bilobatum. Se destaca, además la abundancia de Desmodus rotundus en este cultivo. Para el bosque deciduo coincidencialmente estas especies también están reportada como abundantes, encontrándose aquí sin embargo una alta proporción de Sturnira lilium y Glossophaga soricina a diferencia del cacaotal. Para estos ambientes se reporta un diversidad alfa (riqueza de especies) de 12,92 para el cacaotal, seguida de 9,45 para el bosque seco y 3,9 para el bosque ribereño en orden decreciente respectivamente estos valores indican la relación entre el número total individuos entre el número de especies por hábitats. Los valores de diversidad alfa obtenidos mediante el calculo del índice de Simpson, estos valores son 0,160559; 0,164773; 0,456812 para el hábitats de cacao, bosque seco y bosque ribereño respectivamente. También se observan los valores obtenidos del índice de Shannon - Wierner: 4, 43 para el cacaotal, 2,28 para el bosque seco y 1,53 para el bosque ribereño respectivamente. Posteriormente se muestran los valores de equidad de Pielou: 0,69 para el cacaotal; 0,71 para el bosque seco y 0,46 para el bosque ribereño respectivamente
Se destaca como abundantes las especies que Medellín et al. (2000). señala podrían ser indicadoras de disturbancia, estas especies coincidencialmente son las mismas en nuestro estudio. Carollia brevicauda, Carollia perspicillata, Artibeus jamaicensis y Sturnira lilium. Esta abundancia esta explicada por los hábitos generalitas de estas especies. Carollia es una especie que prefiere alimentarse de frutos asociados a especies vegetales sucesionales que conforman los bosques que protegen al cacao o están presentes en los bosque seco donde abundan plantas de Cecropia sp. y Piper sp. Nosotros encontramos que Carollia perspicillata es la especie más abundante,tanto en las plantaciones de cacao como en el bosque seco lo cual refuerza la idea de que estos cultivos constituyen hábitat muy intervenidos y la distribución de estas especies en el bosque seco al igual que en asociadas en el bosque de cacao, tal vez se hayan afectados por la vecindad de estas plantaciones. La diversidad beta encontramos que 28 especies están presentes tanto en el cacaotal como en el bosque ribereño, 19 especies en el bosque ribereño y el bosque deciduo y 23 especies en el cacaotal y el bosque deciduo, encontrándose Los valor en entre estos dos hábitats posiblemente debido a la vecindad entre estos dos ambiente, su proximidad espacial es aprovechada por los murciélagos para explotar ambos ambientes simultáneamente. La diversidad Beta se calcula a partir de la diversidad alfa promedio, al igual que los valores de complementariedad y el índice de Jaccard para todos los hábitats(Molinari, J. 1989). Se muestran los valores del índice de Whittaker para las relaciones de hábitats: cacaotal-bosque ribereño; bosque ribereño-bosque seco; bosque seco-cacaotal. La diversidad gamma fue calculada a partir del índice de Simpson y un valor relativo de alfa que considera los valores de diversidad alfa de los tres hábitats, este índice también lleva implícita la consideración de la proporción de hábitats en función del área que ocupan dentro del parque, dicha medida no es exacta, por lo cual este valor se considera sesgado y no indica una medida exacta de diversidad. La diversidad gamma encontrada para todos los ambientes fue de 0,1363.
Según Tilman (1999), los estudios de Biodiversidad enfrentan varias barreras logísticas cuando se miden parámetros como numero de especies y endemismos principalmente de naturaleza logística, estos patrones que describen los factores ecológicos que se podían correlacionar con una diversidad alta de especies, llegando a la conclusión que la diversidad aumentaba con la cantidad y equitabilidad de la precipitación y en menor grado con la fertilidad del suelo. En el cultivo de cacao (Theobroma cacao) esta variable no es tomada en cuenta, pero sin embargo se puede afirmar que para este cultivo y para los otros ambientes de nuestro estudio, un suelo pobre, no es una variable que pudiera determinar la presencia o la ausencia de pequeños mamíferos. Las especies de murciélagos se distribuyen, normalmente, según jerarquías de abundancia desde algunas especies muy abundantes hasta algunas muy raras. Generalmente, en las comunidades de mamíferos lo normal es que haya bastantes especies raras, pocas especies abundantes y muchas especies con una abundancia intermedia. Es muy probable que este tipo de relación especie /abundancia, se dé siempre que los elementos del sistema interaccionan y compiten por unos recursos limitados. Medellín et al. (2000) señala que los murciélagos son una herramienta importante por estar presentes en diversos niveles tróficos, son especies ricas y abundantes, ecológicamente diversas, fáciles de muestrear y de responder a cambios ambientales. Esta claro que podemos usar los murciélagos como indicadores de hábitats disturbados pero hay que tener en cuenta los factores relacionados con el indicador en sí mismo, los recursos asociados, su micro hábitat y sus relaciones con otras especies (Pearson 1994).
En los hábitats muestreados correspondientes a áreas de cultivo (Theobroma cacao) existen relaciones definidas entre el número de especies encontradas y la extensión del hábitat, este hábitat se renueva constantemente y su vegetación nunca podría ser considerada como original ya que es alterada continuamente a lo largo del año, cuando se dan las podas y se cosecha. Sin embargo esto ocurre a pequeña escala, es importante, además, destacar la importancia de la cercanía, proximidad (o adyacencia) de hábitats (bosque seco) que de alguna manera influencian en la diversidad y con la cual se establecen importantes interacciones ecológicas entre las especies consideradas transitorias. De hecho hemos encontrado una complementariedad entre el cacaotal y el bosque ribereño de un 0,09 como valor mas bajo, el valor más alto es el del bosque ribereño con el bosque seco 0,38 estos nos indica mayor diferencia entre ambos hábitats. Mientras el valor este más cerca este de 0, los hábitats comparados son más similares o más complementarios con respecto a la composición de especies, es decir el bosque de cacao es similar en composición de especies al bosque ribereño. No fue así, sin embargo con los valores obtenidos cuando comparamos el cacaotal con el bosque seco, en este caso el valor fue de 0,36 y el valor más alto 0,38 para bosque ribereño-bosque seco nos indica que el más amplio rango de diferencia fue el que se establece entre estos ambientes. En cuanto a las plantaciones de cacao como parte del bosque ribereño donde este cultivo pasa a sustituir al segundo estrato arbóreo, en especial en aquellas plantaciones a las que se le aplica un mantenimiento continuo, tenemos que el bosque ribereño ha sustituido al bosque original, formando una mezcla o bosque mixto entre lo natural y el cultivo de cacao dando como resultado, desde la perspectiva florística, un bosque ribereño modificado con la presencia de plantas de cacao, nos atrevemos a sugerir un nombre para este tipo de bosque presente en la del Estado Aragua ubicada dentro de la Costa Central de Venezuela, el cual denominamos "Bosque Ribereño de Cacao".
La importancia del conocimiento de la estructura de estos bosques, radica en la valoración de los mismos para la conservación de la diversidad de vertebrados (Visbal et al. 1992). Hector et al. (2001) señala que en muchos casos, los argumentos para la conservación están basados en procesos funcionales del ecosistema, sin tomar en cuenta las especies que en muchos casos, constituyen indicadores más fuertes para la toma de decisiones a la hora de conservar, sin embargo existe un nuevo movimiento científico que busca argumentos más fuertes basados en la acumulación experimental de evidencias para integrar el abstracto concepto de biodiversidad con el de funcionalidad del ecosistema, sin embargo. recomendamos para el futuro de las mediciones de diversidad debe ir encausado mas hacia medir las características de las comunidades y su ambiente, y no solo el numero de especies capturadas por ambiente. Un estudio similar realizado por Parra (2004), para cultivos de cacao señala la importancia de estos y su relación con la composición faunística de aves, indican que esta depende en gran parte de la sombra que los árboles del dosel proveen y de la densidad de la cobertura vegetal. En este sentido Parra (2004) señala que los cultivos con árboles de sombra como el cacao y el café se vislumbran como la esperanza para la conservación de la diversidad biológica, ante los aumentos proporcionales de la deforestación, ya que, la tendencia actual implica la sustitución del cacao de sombra, por plantaciones de sol, lo cual implicaría una disminución en la riqueza faunística que estos bosques albergan. El cacaotal constituye un oasis para muchas especies de aves que se ven atraídas por el suministro continuo de agua alimento y refugio, recursos escasos en ambientes circunvecinos durante esa época del año. Nuestros resultados, nos llevan a señalar que la estructura florística asociada a los cacaotales de sombra, dentro de hábitats muy fragmentados como lo son los bosques tropicales, proveen de recursos ecológicos, alimentarios y de refugio a numerosas especies, de allí que estos albergan una amplia diversidad de fauna de vertebrados e invertebrados, los cuales se benefician de los microhabitats que allí encuentran.
Conclusiones
La riqueza de especies de las comunidades de quirópteros fue determinada usando el índice de diversidad de Margalef, el valor mas alto corresponde a las especies asociadas a una plantación de cacao (Theobroma cacao), que en este estudio hemos denominado bosque ribereño de cacao, los valores para el bosque seco y el bosque ribereño siguen en orden decreciente respectivamente. Para la diversidad beta de las comunidades de quirópteros presentes entre una plantación de cacao, un bosque deciduo y en un bosque ribereño, encontramos que el valor más alto es el que se establece entre el bosque del cacao y el bosque seco, y esto se debe posiblemente a su proximidad espacial. Los valores de complementariedad de hábitats indican que el cacaotal es similar al bosque ribereño en cuanto a la composición faunística que estos bosques albergan. Los quirópteros mantienen una relación mutualista con el cultivo de cacao (Theobroma cacao), este le sirve de refugio, es propicia para la captura de las presas de los murciélagos insectívoros, los provee de una gran gama de recursos alimenticios para una comunidad de amplia variedad de hábitos alimenticios. Los murciélagos constituyen el grupo de vertebrados ideal para ser utilizadas como indicadores de diversidad biológica de la vertiente norte del Parque Nacional Henri Pittier, puesto que se mantienen una estrecha relación con la estructura de la vegetación, los recursos bióticos y actividades antropogénicas.
Se recomienda aumentar el número de horas de campo, a fin de disponer de mayor cantidad de datos, para este grupo zoológico con tan amplia distribución y amplia variedad de gremios alimenticios. Recomendamos el uso de detectores de Ultrasonido a fin de identificar la presencia de especies de vuelo libre, que jamás podrán ser colectadas usando el método de mallas de neblina (Willig & Selcer 1989). Estandarizar los protocolos para la realización de inventarios de mastofauna, con el fin de que estos puedan ser utilizados de manera universal por otros investigadores y estos datos sean amoldables a la aplicación de cualquier modelo estadístico (Zar 1999). Ampliar las investigaciones con referencia a cultivos tradicionales (cacao, café, etc.) a fin de entender las dinámicas de estas poblaciones vegetales y sus relaciones con la fauna asociada a estos cultivos. Establecer políticas de conservación de los cultivos tradicionales de cacao dentro del Parque Nacional Henri Pittier, entendiéndose estos como reservorios importantes de biodiversidad.
Agradecimientos
Al Instituto de Zoología Agrícola de la Facultad de Agronomía, Universidad Central de Venezuela. Al Dr. Javier Sánchez y al Museo de la Estación Biológica "Rancho Grande", al apoyo incondicional del Ing. Andrej Skora, a Stefán Skora por su especial ayuda en el campo, a Kay Rosemberg y toda la gente de la Hacienda "La Sabaneta" en Choroní, gracias infinitas y a FUNDACITE Aragua por el financiamiento parcial de esta investigación.
Referencias
Aguilera, M. A. Azocar & E. Gonzalez-Jimenez. 2002. La Biodiversidad en Venezuela. Fundación Polar. Ministerio de Ciencia y Tecnología. Fondo Nacional de Investigaciones Científicas y Tecnológicas. 1080 pp.
Anzoategui, P. 1994. Cambios en el uso de la Tierra en la Parroquia Choroní, Estado Aragua, Venezuela. Universidad Central de Venezuela, Facultad de Agronomía, Trabajo de grado para optar al titulo de Ingeniero Agrónomo. 240 pp.
Ezcurra, E. 1990. ¿Porqué hay tantas especies raras? La riqueza y rareza biológicas en las comunidades naturales. Ciencias (No. especial) 4: 82-88.
Fernandez-Badillo, A., R. Guerrero, R. LordO, J. Ochoa & G. Ulloa. 1988. Mamíferos de Venezuela: Lista y Clave para su identificación. Editado por ASOVEN y MIZA-UCV.185 pp.
Fernandez-Badillo, A. 2000. El Parque Nacional Henri Pittier. Revista de la Facultad de Agronomía. Alcance Nº 60 Universidad Central de Venezuela. 284 pp.
Fleming, T., E. Hopper & D. Wilson. 1972. Three Central American bat communities: structure reproductive cycles and movement patterns. Ecology 53(4) 554-569.
Fox, B.J. & M.D. Fox. 2000. Factors determining mammal species richness on habitat islands and isolates: habitat diversity, disturbance, species interactions and guild assembly rules. Global Ecology & Biogeography 9: 19-37.
Gaston, K. & J. Spicer. 1988. Biodiversity: an introduction. Blackwell Science Ltd. Editorial Office: Osney Mead Oxford OX2 OEL.113 pp.
Halffer, G. & E. Ezcurra. 1992. ¿Qué es la Biodiversidad? Acta Zoológica Mexicana. Vol. Especial-La Diversidad Biológica de Ibero América.
Hector, A., J. Joshi, S.P. Lawler, E.M. Shehn & A. Wilby. 2001.Conservation implications of the link between biodiversity and ecosystem functioning. Oecologia 129: 624-628.
Holdridge, L.R. 1978.Ecología de las zonas de vida. Organización de Estados Americanos (OEA). Instituto Interamericano de Ciencias Agrícolas, San José, Costa Rica. 123 pp.
Hurlbert, S.H. 1978. The non-concept of species diversity: a critique and alternative parameters. Ecology 59: 67-77.
Linares, O. 1998. Mamíferos de Venezuela. Editorial Sociedad Conservacionista Audubon de Venezuela. Caracas.692 pp.
Manama, D. & Rievero, G. 1994. La ocupación espacial en la parroquia Choroní (Estado Aragua) y sus implicaciones Ambientales en el Parque Nacional Henry Pittier. Universidad Central de Venezuela, Facultad de Humanidades y Educación, Escuela de Geografía. Trabajo especial de Grado. 183 pp.
Magurran, A. 1988. Ecological diversity and its measurement. Princeton University Press. 179 pp.
Medellin, R. & M. Equihua. 1998. Mammal species richness and habitat use in rainforest and abandoned agricultural fields in Chiapas, Mexico. Journal of Applied Ecology 35: 13-23.
Medellin, R., M. Equihua & M. Amin. 2000. Bat diversity and abundance as indicators of disturbance in Neotropical rainforests. Conservation Biology 16(6): 1666-1675.
Molinari, J. 1989. La diversidad ecológica: un enfoque unificado, conceptual y metodológico para su cuantificación. Universidad Central de Venezuela. Trabajo de grado presentado para optar al Titulo de Doctor en Ciencias mención Ecología. Facultad de Ciencias, 260 pp.
Moreno, C. 2000. Métodos para medir la biodiversidad. Manuales & Tesis SEA. UNESCO, CYTED. España. 83 pp.
Moreno, C. & G. Halffer. 2000. Assessing the completeness of bat diversity inventories using species accumulation curves. Journal of Applied Ecology 37: 149-158.
Moreno, C. & G. Halffer. 2001. Spatial and temporal analysis of , and diversities of bats in fragmented landscape. Biodiversity and Conservation 10: 367-382.
Noss, R.F. 1990. Indicator for monitoring biodiversity: a hierarchical approach. Conservation Biology 4(4): 355-365.
Parra, L. 2004. Estructura de la comunidad de aves del sotobosque de un cultivo de cacao (Theobroma cacao) en el valle del Municipio Costa de Oro, Estado Aragua, Norte de Venezuela. Universidad Central de Venezuela. Trabajo de Grado presentado para optar al titulo de Magíster Scientarum en Zoología Agrícola. 65 pp.
Pearson, D. 1994. Selecting indicator taxa for the quantitative assessment of biodiversity. Phil. Trans. R. Soc. Lond .B 345: 75 -79.
Rojas, C. 1994. Murciélagos de un bosque ribereño de la vertiente norte del Parque Nacional Henri Pittier, Aragua, Venezuela. Universidad Central de Venezuela, Facultad de Agronomía, Trabajo de grado para optar al titulo de Ingeniero Agrónomo, 140 pp.
Ortiz, I. 1993. Evaluación de la entomofauna asociada al fruto del cacao (Theobroma cacao) en Ocumare de la Costa, Estado Aragua, Venezuela. Universidad central de Venezuela, Facultad de Agronomía, Trabajo de grado para optar al titulo de ingeniero Agrónomo, 140 pp.
Schulze, M., N. Seavy & D. Whitacre. 2000. A comparison of the Phyllostomid bat assemblages in undisturbed neotropical forest and in forest fragments of slash and burn farming mosaic in Petén, Guatemala. Biotropica 32(1): 174 -184.
Sokal, R. & F. Rohlf. 2000. Biometry. 3° Edicion Freeman. San Francisco.
Solbrig, O. 1991. Biodiversity scientific issues and collaborative research proposal. MAB Digest N° 9 UNESCO. Paris. Francia.
Tilman, D. 1999. The ecological consequences of changes in biodiversity: a search for general principles. Ecology 80(5): 1455 -1474.
Varela, L. 1999. Diagnostico de la producción del cacao en el Estado Aragua. Informe para Fundacite Aragua (Agenda Cacao) 71 pp.
Visbal, R., J. Manzanilla & A. Fernandez. 1992. Importancia de los vertebrados del Parque Nacional Henri Pittier y consideraciones para su conservación. IMPARQUES 73 pp.
Willig, M. 1986. Bat community structure in South America a tenacious Chimera. Revista Chilena de Historia Natural 56: 151 -168.
Willig, M. & K. Selcer, K. 1989. Bat species density gradients in the New World: statistical assessment. Journal of Biogeography 16: 189-195.
Zar, J. 1999. Biostatistical Analysis. 4° Edition Prentice Hall. New Jersey.