


 Email this Article
Email this Article Print
Print Download (PDF)
Download (PDF)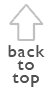
Abstract 
A comparison of vegetation changes during and after EN 1997/98 at a coastal desert site and a dry woodland environment 90 km inland is given. Initially, both sites react similarly to El Niño: short-lived herbs and grasses sprout quickly. But the ongoing succession and retrogression differs in the desert and the woodland due to differences in precipitation at the two sites as well as differences in disturbance regimes (fire and burrowers) and in the intensity of land use. Results are discussed in the context of global climate change and are applied to human land use issues where ruderals and nonindigenous species invade and change species composition. It is hypothesized that increased ENSO frequency and intensity create greater phytomass that in turn encourages farmers to graze more cattle which leads to degradation with a shift from dry forest to shrub savanna.
Introduction
Taking northwestern Perú as a core region of
ENSO
This
study examines ecosystem dynamics after El Niño events in
northwestern Perú. The Sechura Desert and the adjacent dry
forest zone of the coastal plain in northwestern Perú (Fig. 1)
have episodic severe flood events and extreme positive rainfall
anomalies. Due to El Niño Southern Oscillation (ENSO), the
study area is marked by the highest variability in annual
precipitation in South America and likely even on earth (Caviedes
1998, Goldberg et al. 1987). During so called "Super-Niños"
the deviation from average precipitation in non-ENSO-years may
surpass 9000 %, such as observed at Talara in 1997/98. Although the
Sechura Desert is the central "wet spot" of ENSO
activity, thorough physiological or ecological research on pertinent
ecosystem responses is just beginning.The original aim of the study was to document plant succession and retrogression during and after El Niño 1997/98, thus seven field sites were chosen crossing the coastal plain along a transect between the coast of Paita and the hinterland near Chulucanas (Fig. 1). The study area is located on the coastal plain in a flat environment lower than 200 m.a.s.l. It includes several granitic and phyllitic outcrops up to 385 m.a.s.l. but is predominantly made up of Pleistocene alluvial deposits and Holocene fossil sand dunes (Fig. 1). In this paper, two sites are examined in detail, one a desert ecosystem (near the coast), the other a dry woodland (inland). We find these sites most typical of the 90 km transect between Paita and Chulucanas. Field observation has continued for more than eight years, i.e., from the pre-Niño phase in October 1997 through April 2005. Previous results (Block and Richter 2000) are combined with ideas of a higher frequency or intensity of El Niño discussed in the context of climate change (Bendix et al. 2000), which raises questions regarding future ecological development in the region. For an extended version of the paper in hand see Richter and Ise (2005).
The non-ENSO average precipitation between Paita and Chulucanas lies between 15 mm/yr in the west and 320 mm/yr in the east. During the last "Super-ENSO" the precipitation gradient ran from around 800 mm/yr at the coast up to 4,000 mm/yr in the preandean foreland (Fig. 1). Usually these high intensity events occur over a limited time period, between December and May (as observed during the 1997/98 event). Thus, during these episodes, floristic elements of another vegetation-zone should be capable of entering and establishing in the desert environment for limited periods and suggests that a fifty-fold increase in precipitation - as was the case in Paita - should cause greater variations in plant cover than the twelve-fold increase that occured in Chulucanas.
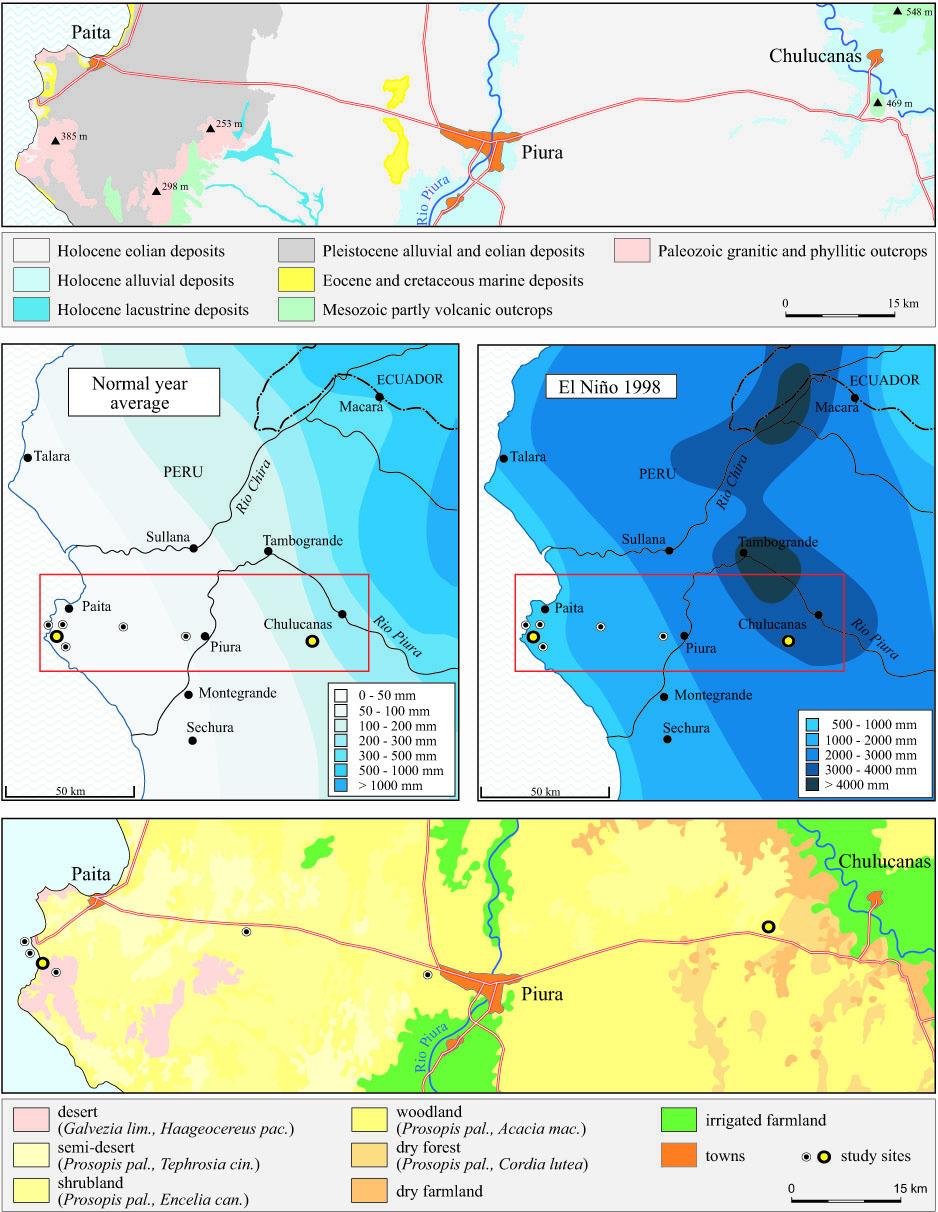
Fig.
1. Geological units in northwestern Peru (above, based on maps of
INGEMMET 1984-1989), rainfall patterns during a non-ENSO year and in
1998 (center, based on data from Proyecto Chira y Piura), and
distribution of vegetation communities (below, based on maps of
INRENA 2000, and on own observations).
Under normal conditions the regional plant formations reflect a gradient between a tropical desert and dry forest. Detailed vegetation maps derived by INRENA (2000) from remote sensing analyses show a patchwork of different communities. Fig. 1 indicates patterns influenced by varying climatic, hydro-geological, and human impacts. In the areas little changed by people, a general feature of all plant formations is the constant presence of Prosopis pallida ("algarrobo") and of Capparis scabrida ("zapote").
Methods
The
two study sites presented here are situated at the two extreme
positions of this climatic and vegetation gradient (fig. 1). Each
study site comprises five sample plots of 400 m² each (20 x 20
m). Since October 1997 field investigations occur twice a year in
March/April and September/October, i.e. at the end of the wet season
(when one exists) and during the second half of the 9- to 11-month
long dry season. The coastal study site (Paita) is located 800 to
1,000 m from the coastline on a sandy plain at 65 m.a.s.l. It sits on
a pediment that descends coastward from the foothills of the Cerros
del Diablo southeast of Paita. Aeolian processes have caused a
residual concentration of gravel on the slightly encrusted surface
("eluvial sserir"). Under the topsoil lies a
loosely-packed sand stratum. The second study site (Chulucanas) is
located at 175 m.a.s.l. in a fossil sand dune area covered by
shrubland and open woodland. While human impact at the Paita site is
negligible, goats, and to minor extent brahma-cattle, cause
continuous grazing pressure at Chulucanas.The sample plots are situated along a N-S-transect at a 20 m interval. Regardless of the plant formation type, plot size is held constant. During each field visit, plant distribution is mapped at one plot at each site. At all plots species composition, abundance and percentage coverage as well as the degree of vitality are taken; life-form for each species is derived from visual inspection while data on habitat-preferences are based on Brako a. Zarucchi (1993) and on Sagástegui Alva a. Leiva Gonzales (1993). Statistical analysis includes the calculation of mean species coverage at each site and the mean sum of coverage for life forms and habitat preferences. Moreover, an analysis of temporal similarity has been applied using root transformed values of mean species coverage. Similarity is calculated with the Euclidean distance measure, and clustered by Wards method.
Results and Discussion
In
the introduction we hypothesize that floristic elements of another
vegetation-zone should be capable of entering and establishing in the
desert environment for limited periods and that the fifty-fold
increase in precipitation - as was the case in Paita - should cause
greater variations in plant cover than the twelve-fold increase that
occurred in Chulucanas. We conclude now,
that this is true for the initial phase after El Niño. In the
following phase, however, successive and retrogressive trends are
more complex. This is displayed by the coverage values shown by the
diagrams of Fig. 2.In the case of the Paita site, the development is restricted mainly to the outstanding peak due to the ENSO rainfall-boom (fig. 2 left). Here, water deficits prevail during the entire post-Niño phase with a short monsoon period in 2001 causing a slight temporary increase in plant cover. Even less effective is a stronger monsoon or Niño-like event in 2002 since several spotfires reduced the effect of the higher water input during March and April. In contrast, at the Chulucanas site the uneven heights of the bars (Fig. 2, right) reflect oscillating trends in plant growth. In this case, the monsoon rainfall events "reset the clock" in terms of refreshing plant development. Thus, different development trends in the desert and in the dry woodland are evident.
At Paita, the short-term boost causes a considerable deviation from the poor coverage during the dry conditions of non-ENSO periods (Fig. 2, left). Therophytic herbs such as Crotalaria, Desmodium, and Exodeconus are the first prominent participants of a micro-succession which starts within one month after the first rains (Fig. 2 a, left). Annual grasses of Aristida and the perennual herbs Alternanthera, Tephrosia, and Tiquilia follow in the next few weeks, showing a longer survival rate. While the quasi-permanent non-ENSO tree-community of Prosopis pallida and Capparis scabrida consists mainly of representatives of the dry forest habitats, the associated herbs and grasses of the short-term ENSO-community originate from desert habitats or are considered native ruderals (Fig. 2b, left). Only during the outstanding peak coverage within the El Niño boost, are taxonomic and structural compositions marked by a strong divergence from the usual. The dendrogram in Fig. 2c (left) reveals a fast assimilation of the post-Niño-community back towards the pre-Niño-situation as a crucial feature for plant successions in the Paita area. Parallel investigations on sandy and rocky sites corroborate this fundamental result for all desert environments, where no effective long-term community change can be expected after Super-Niños. Nevertheless, there are permanent structural vegetation changes enforced by primary production of woody species as a reaction to heavy rainfalls creating renewed ground- and porewater reservoirs and overall higher moisture availability.
The dendrogram of Chulucanas displays a much more vivid vegetation change than that of Paita (Fig. 2c). The quasi-permanent non-ENSO woody community consists of indigenous species such as Prosopis pallida, Capparis scabrida, Acacia macranthera and Cordia lutea. In contrast, the episodically promoted short-term herb and grass layers of ENSO, monsoon, and post-fire events in the inland are dominated by a varying composition of many non-indigenous ruderals and a lower number of native sand species. The dendrogram shows up four mayor groups: The first one (left) is characterized by low coverage values. It includes most samples from the dry seasons (each October in 1997-2001) and from April 1999, when plant cover was still low due to a severe fire event in Sept. 1998. The second group is joined with the first one and comprises the last phase of development, i.e. the remaining dry community including a final enlargement of Acacia-shrubs (from October 2003 to now). Its propagation was first hampered by the root competition among herbs, grasses, and shrub shoots within the upper rhizophere. After an initial activation of a plentiful seed bank, relatively few individuals of Acacia survived up to a point of abruptly augmenting growth rates. This point coincides with the state when roots reached deep soil moisture. We assume that after this "break-through" a high survival rate is the case, even through a future long-term desiccation. The third group is less similar; this unit is marked by a boost-phase of Ipomoea incarnata and Tribulus terrestris (April 2000 and 2001) which are the most apparent members of the post-fire development sprouting from rhizomes or tubers. The last group shows an outstanding dissimilarity to the other groups. It consists of post-rainfall associated plants and indicates that in 2002 the effects of the second monsoon year are comparable to those of the Niño event in April 1998. In these stages, hemicryptophytic grasses such as Anthephora hermaphrodita and Cenchrus pilosus and some herbaceous climbers make up an important food supply for livestock.
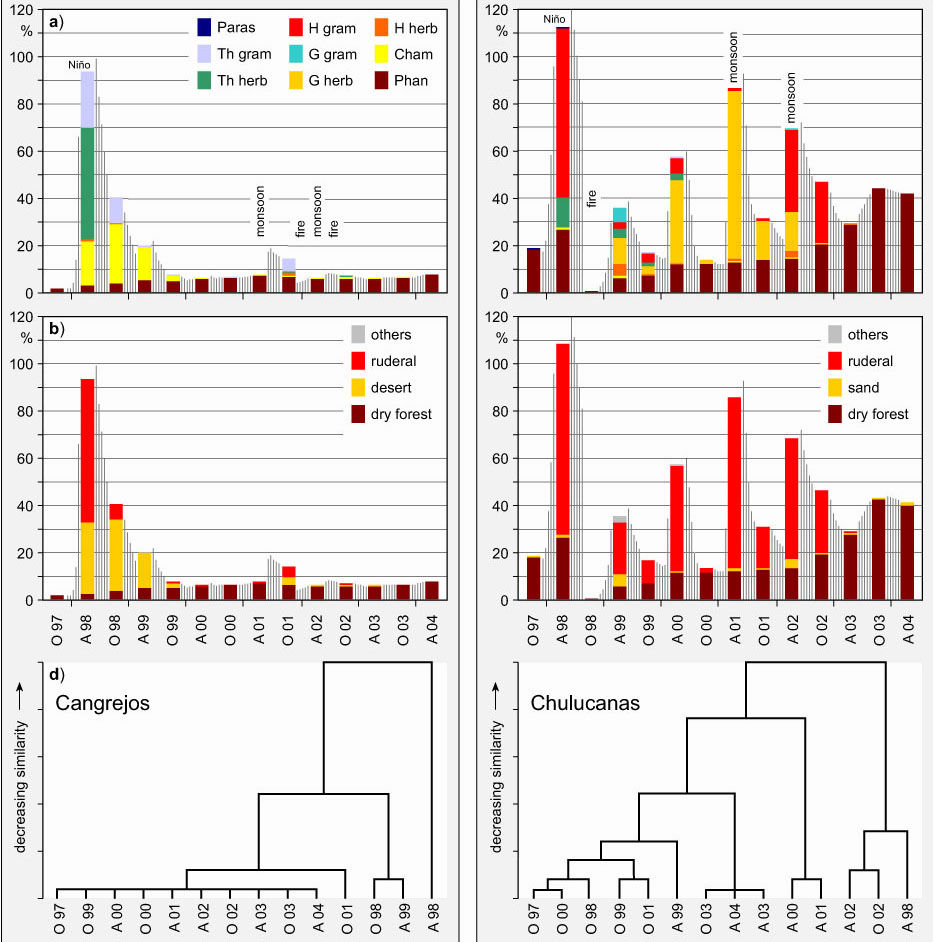
Fig. 2: Development of life-forms (a), habitat preferences (b) as well as
floristic similarity in time (c) between 1997 and 2004 at the Paita
and Chulucanas sites.
In Paita, variations in plant coverage between greater and lesser rainy seasons are hardly notable, while the Chulucanas site displays a significant dependence on the more variable rainfalls. Furthermore, in Paita, therophytes and slightly lignified chamaephytes play a decisive role restricted to the first part of succession, whereas the life-form spectra of Chulucanas are characterized by longer lived hemicryptophytes, geophytes and micro-phanerophytes. Native elements rule the floristic structure of the plots at Paita, while more widespread and invasive ruderals govern the ecosystems in Chulucanas. This results from the differing human impact intensity: grazing as well as irrigated field cultivation in the hinterland promotes the input and propagation of wind- and animal-dispersed (i.e. exozoochorous) weeds, some of them neglected by cattle; at the coast grazing is minimal and there are no fields. The response to Super-Niño 1998 and the two monsoon events lead to an increasing distribution of shrubs and trees. We see this as a savannification process. However, at the same time overgrazing may lead to a degradation of the floristic composition.
Climate change and vegetation development: dangers and
opportunities
For
5,000 years of record, various ENSO proxy indices (marine sediments,
ice-cores, ocean settings, tree-rings etc.) carry evidence of the El
Niño phenomenon. Sand deposits in
the catchments of the Rios Piura (fig. 1) and Chira seem to result
from strong Niño-phases between 3,900 and 3,600 BP as well as
between 2,800 and 2,500 BP (Martin et al. 1993). Furthermore, fluvial
accumulations of the Rio Casma indicate a return rate of around 1,000
years for "Mega-Niño events" which by far exceeded
the intensity of recent Super-Niños (Wells 1990). Thus
irregular Niño-frequencies and -intensities seem to be a
normal natural phenomenon.
Results from high-resolution ocean-atmosphere models presented by Timmermann et al. (1999) show, that the recent Niño-phase must not be judged as normal, but rather as a superposed low-frequency trend within a climate change that will be accelerated over the coming decades. By coupling the model with a rise of man-made CO2-emissions the authors Latif (1999) supposes an increase of SST of >3°K for the eastern tropical pacific region up to the year 2100 (fig. 3). In this context an greater probability of a higher intensity and frequency of Niño-like weather events, including the monsoon effects, becomes realistic.
Fig. 3: Indicator values for past and future El Niño activity (b):
Southern oscillation index (SOI) between 1882 and now (source: http://sevilleta.unm.edu/data/contents/SEV016/, http://sevilleta.unm.edu/data/contents),
development of sea surface temperatures (SST) between 1860 and now
and scenario of a future increase based on coupled ocean-atmosphere
models (source: Latif 1999) and annual rainfall amounts at Piura from
1932 until now (Data: Proyecto Chira y Piura). Possible future
development of vegetation before, during and after Super-Niños
without human impact on a desert site (a) and with overgrazing impact
in a dry forest site (c) is shown.
The precipitation record available for Piura starts in 1932 (Fig. 3). A phase of relatively high precipitation activity has been recorded between 1930 and 1945 followed by a drier phase until 1973. With the Super-Niño of 1982/83 a phase of increased Niño frequency and greater intensity begins, including a smaller event in 1986/87 as well as the double-Niño 1991/93, and culminating in the recent 1997/98 phenomenon (Bendix et al. 2000). The same trend is found in the much longer chronology of El Niño indices presented by Quinn et al. 1987.
Of course, it is speculative to link a single ENSO-event with climate change and hence, with ecosystem change. Nevertheless, the attempt to illuminate possible post-Niño vegetation scenarios is important, as the actual knowledge provides appropriate ideas. A more frequent disturbance regime obviously shortens the cycles of a system and offers more possibilities for pioneer species to occur. Consequently, a possible future rapid change in weather phenomena supporting boost effects must accelerate the input of its most dominant species including an explosive enrichment of invasive ruderals.
An advanced interpretation might be achieved by revealing to what extent the "vegetation of the moment" may advance under more frequent Niño- and Niño-like monsoon events (Fig 3). In the case of no human impact on vegetation the most recent relevés at Paita give hints for future development. For example, Alternanthera halimifolia and Tephrosia cynerea had negligible abundance in the study plots up to the last Super-Niño. However, most recent observations display a tendency of possible survival of these species if the frequency of El Niño-cases or unforeseen moist-warm incidences will increase. Shrubs such as Galvezia limensis and Waltheria ovata in the Sechura Desert or Acacia macracantha and Encelia canescens in the dry forests of Chulucanas are indicating a progression as well. Further propagation of the most abundant trees Prosopis pallida and Capparis scabrida is obvious. However, in natural vegetation, droughts, fires, storms and floods will not cause floral changes if ruderal seed pools are not available.
In terrains with grazing and an increase of farming typical for the inland regions, diverse dynamic characteristics and strategies of invading ruderals such as cohort senescence, toxic effects and competitive or regenerative qualities must be considered. Recent post-fire expansion of Acacia macracantha and also of Cordia lutea leads to a further propagation of the shrubby members which might hamper the two dominant tree species by root competition. A progressive invasion of additional annuals may lead to a provisional release of new satellite species while non-native perennial herbs may increase the total vegetation cover (Fig. 3). Later on, whether there will be a collapse or a renewal of dominant taxa, a decline of native herbs must occur as an inevitable temporary fact: At least in transitory phases, plant selection by grazing as well as by specific inhibitory substances, might produce permanent fluctuations in species numbers.
Thus, concerning its effects on vegetation, climate change can not be regarded to be dissociated from global change. According to Beerling (1995) human activity in combination with an increased frequency of extreme events allows species invasions and homogenizes species composition on a zonal scale. "Phytoglobalization", i.e. an accelerated mobility of invaders by human impact, is triggered by and connected with the increasing economic trade and mobility of people. Extreme weather situations due to a change in circulation patterns may cause new types of disturbances or new magnitudes of boost effects, thus giving space for plant invasions. Whether equivalent processes in northwestern Perú must be considered dangerous depends on one's personal opinion - at least invaders do not seem to threaten the desert ecosystems in a substantial way.
However, a real problem is the decline of plant productivity and land degradation resulting from overgrazing because El Niño rains allow peasants to expand extensive goat breeding. In principal, the peasants are right since many land-use experts and ecologists are beginning to emphasize the positives effects of El Niño, for example now discussing a "window of opportunity for the restoration of degraded arid ecosystems" (Holmgren a. Scheffer 2001). Thus, forecasting ENSO episodes is employed for semiarid rangeland and production systems to identify the risk of overgrazing and fire during dry years and to stimulate pasture and woodland regeneration during and after rainy years.
Enhanced productivity from El Niño precipitation contrasts with the widespread effects of desertification. Although being only a temporal event, a recovery during the post-Niño phase may be based on the resilience of the still existing woodlands. The long-term effects of a brief regeneration cycle offer a worthwhile situation to use rainfall in the most efficient way. Consequently, the three to four years following a strong El Niño event is a period of highest importance and enhanced necessity to support and control the regeneration processes which are driven by the energy flows within the terrestrial ecosystems in northwestern Perú.
References
Beerling, D.J. 1995. General aspects of plant invasions: an overview. In: Pyek, P., M. Prach, M. Rejmánek & M. Wade (eds.): Plant invasions. General aspects and special problems. Amsterdam
Bendix, J., & Bendix & M. Richter. 2000. El Niño 1997/1998 in Nordperu: Anzeichen eines Ökosystem-Wandels? Petermanns Geographische Mitteilungen 144: 20-31.
Block, M. & M. Richter. 2000. Impacts of heavy rainfalls in El Niño 1997/98 on the vegetation of Sechura Desert in Northern Peru. Phytocoenologia 30: 491-517.
Brako, L. & J.L. Zarucchi. 1993. Catalogue of the Flowering Plants and Gymnosperms of Peru. Monographs in Systematic Botany from the Missouri Botanical Garden 45. St. Louis.
Caviedes, C.N. 1998. Influencia de ENSO sobre las variaciones interanuales de ciertos rios en America del Sur. Bull. de l´Intsitut Français d´Etudes Andines 27: 627-642.
Goldberg, R. & G. Tisnado & R. Scofield. 1987. Characteristics of extreme rainfall events in NW Peru during the 1982/83 El Niño period. - Journal of Geophysical Research 92: 14225-14241.
Holmgren, M. & M. Scheffer. 2001. El Niño as a window of opportunity for the restoration of degraded arid ecosystems. Ecosystems 4. 151-159.
INGEMMET 1984-1989. Mápa Geológico del Perú 1:100.000. Lima.
INRENA-Proyecto algarrobo. 2000. Mapa de bosques secos del Departamento de Piur& Memoria descriptiva y mapa (second edition). Piura.
Martin, L.,M. Fournier, P. Mourguiart, A. Sifeddine, B. Turco, M.L. Absy & J.-M. Flexor 1993. Southern oscillation signal in South American palaeoclimatic data of the last 7.000 years. Quaternary Research 39: 338-346.
Quinn, W.H., V.T. Neal & S.E. Mayolo de. 1987. El Niño occurrences over the past four and half centuries. Journal of Geophysical Research 92: 14449-14461.
Richter, M. & M. Ise. 2005. Monitoring plant development after El Niño 1997/98 in Northwestern Peru. Erdkunde 59(2) : 136-155
Sagástegui Alva, A. & S. Leiva Gonzales. 1993. Flora invasora de los cultivos del Peru. Trujillo.
Timmermann, A., J. Oberhuber, A. Bacher, M. Esch, M. Latif & E. Röckner. 1999. Increased El Niño frequency in a climate model forced by future greenhouse warming. Nature 398: 694-697.
Wells, L.E. 1990. Holocene history of the El Niño phenomenon as recorded in flood sediments of northern coastal Peru. Geology 18: 1134-1137.